Adaptation of Photosynthesis : Parametrisation from Laboratory Experiments
|
Résumé du projet : The photoadaptation process in autotrophic phytoplankton optimizes inorganic carbon fixation in suboptimal conditions of photon flux and nutrient supply. Interestingly, the variations and concomitance in spatio-temporal limiting factors comprise a ecological reality which is rarely considered in model systems and in laboratory experiments. This project will unite diverse inputs from physicists, system modelers, and biologists who will attempt to reproduce experimentally the complexity of the natural environment based on analyses of preliminary hydrodynamic and biological data obtained from campaigns such as POMME. New experimental data will then consolidate the conception of photoadaptation models. The pertinence of these models will be analyzed as a function of the inherent constraints found in modeling physical and biological interactions in pelagic ecosystems. |
||||||||
|
click to go to ....
|
||||||||
I.1.
ETAT DE L'ART - CADRE GENERAL |
||||||||
La photoadaptation est un des processus qui permettent aux cellules phytoplanctoniques autotrophes d'optimiser leur dťveloppement dans un environnement oý les conditions de croissance sont physiologiquement sub-optimales (photoadaptation est ŗ prendre ici dans sa connotation phťnotypique, et non gťnotypique). De son efficacitť dťpend le flux de carbone associť ŗ la production primaire. L'ťtude in situ de ce processus est extrÍmement complexe car il intŤgre l'influence de nombreux facteurs que l'on ne maÓtrise pas, dont le plus ťtudiť a ťtť le flux de photons. La tempťrature et les ťlťments nutritifs, notamment le nitrate, jouent ťgalement un rŰle prťpondťrant dans la rťgulation de la synthŤse (photoadaptation) et de la composition (adaptation chromatique) du pool pigmentaire (Cullen et al. 1993) . MÍme au laboratoire, les ťtudes expťrimentales se sont essentiellement cantonnťes ŗ l'effet de la lumiŤre (Marra 1978a; Marra 1978b) , et les modŤles de photosynthŤse utilisťs ne font gťnťralement intervenir que ce dernier paramŤtre (Cullen et Lewis 1988; Lande et Lewis 1989).
Malgrť cela, la photoadaptation est largement sous-reprťsentťe dans les modŤles de rťseau trophique. Ces derniers sont nťcessairement simplifiťs ŗ cause de considťrations restrictives liťes ŗ la reprťsentation dťterministe de systŤmes complexes (surparamťtrisation, incertitude sur les processus et les paramŤtres, validation incertaine, etc.). Comme la pratique de la modťlisation est rapide comparativement aux avancťes expťrimentales (et moins chŤre aussi), la rťsolution de problŤmes conceptuels complexes a presque toujours ťtť conduite sur l'improvisation de relations empiriques (typiquement loi du minimum, loi multiplicative, etc.), non validťes par l'expťrience, car non ťtudiťes ni testťes (Sciandra et al. 1997) . Les modŤles rťcents qui prennent en compte un couplage structurel des effets simultanťs de l'azote, de la lumiŤre et de la tempťrature sont pour l'essentiel confrontťs ŗ des mesures acquises ŗ l'ťtat stable (Flynn et al. 2001; Geider et al. 1997; Geider et al. 1998; Zonneveld 1998) . Il faut dire que l'approche expťrimentale portant sur des systŤmes multiparamťtriques et instationnaires (Davidson et al. 1999) s'est faiblement dťveloppťe par le passť, compte-tenu des difficultťs inhťrentes ŗ l'ťtude des organismes planctoniques d'une faÁon gťnťrale, et ŗ celles d'exercer un contrŰle complexe, prolongť et intelligent sur ces mÍmes organismes, en particulier. L'apparition d'outils propres au dťveloppement d'automates permet d'envisager maintenant des expťriences d'un autre type, plus adaptťes ŗ l'ťtude des processus non linťaires.
|
||||||||
Ces considťrations d'ordre gťnťral nous incitent ŗ penser que les approches expťrimentales et thťoriques doivent Ítre conduites diffťremment de par le passť pour qu'une rťelle synergie puisse se dťvelopper entre elles. Il est notamment nťcessaire d'ťtudier les processus de photoadaptation sur des organismes replacťs dans un environnement artificiel mais contrŰlť (dont on maÓtrise les paramŤtres), c'est ŗ dire susceptible de reproduire autant que possible les conditions caractťristiques d'un biotope particulier (nature des facteurs limitant, ťchelle de temps caractťristique de leur variation, etc.). Les multiples interactions de type proie-prťdateur existant dans un rťseau trophique conduisent les organismes ŗ rťtroagir sur leur propre environnement (ťpuisement du substrat, auto-ombrage, etc.) et ŗ modifier les facteurs qui affectent leur croissance. Par contre, les phťnomŤnes qui dťterminent le forÁage principal du rťseau trophique et qui n'en subissent quasiment aucune rťtroaction sont d'origine physique (advection, convection, diffusion, ondes internes, etc.). Introduire intelligemment cette composante essentielle et inaliťnable du fonctionnement des ťcosystŤmes dans des expťriences de laboratoire est un rťel problŤme conceptuel qui nťcessite la contribution de physiciens. Un concertation en amont des expťriences de laboratoire avec les modťlisateurs est donc nťcessaire.
Le saut qualitatif que permet ce nouveau type d'expťrimentation dans l'ťtude des processus non linťaires caractťristiques de la photoadaptation n'est possible que si l'on parvient ŗ reproduire un environnement dynamique, multiparamťtrique, continu et instationnaire, en rupture avec les conditions d'expťrimentation traditionnelle oý l'ťtude porte gťnťralement sur une gamme de variation discrŤte d'un paramŤtre stabilisť dans le temps. Ceci implique qu'un effort technique important soit conduit, notamment pour automatiser les contraintes apportťes au systŤme, ainsi que la mesure ŗ haute frťquence de ses variables
L'approche multiparamťtrique est essentielle pour une ťtude approfondie de la photoadaptation dans un environnement oý la tempťrature, les flux de photons et de nitrate interagissent dans la rťgulation de la synthŤse pigmentaire. Ce processus a surtout ťtť ťtudiť en fonction du flux de photons et de la composition spectrale de la lumiŤre dans des conditions de croissance optimale de nutriment et de tempťrature. Pourtant, parce que la synthŤse de pigments et d'enzymes carboxyliques nťcessite une importante mobilisation protťique, la photoadaptation est nťcessairement dťpendante du statut azotť des cellules. Dans quelle mesure ce dernier affecte-t-il leur capacitť ŗ optimiser leur pool de pigment est une question mal ťlucidťe et surtout non prise en compte dans les modŤles (Sciandra et al. 1997) , et qu'il faudra aborder. La production autotrophe rťsulte in fine d'une synthŤse protťique. L'interdťpendance physiologique de la fixation de carbone et de l'assimilation de nutriment inorganique, notamment ŗ travers la rťgulation pigmentaire, fait que la croissance ne peut probablement pas Ítre une fonction simple (loi du minimum ou multiplicative) des actions conjuguťes des flux de photons et de nutrilites. Pour le savoir, il faut reconstituer un environnement oý ces flux sont simultanťment limitants pour la croissance.
En s'adaptant, une cellule de phytoplancton tend, par dťfinition, vers un ťtat d'ťquilibre caractťrisť par un optimum de croissance relatif aux conditions externes de lumiŤre et de nutrilites. Si ces conditions sont elles-mÍmes variables dans le temps, l'efficacitť d'adaptation sera en relation avec le rapport des vitesses d'adaptation des cellules et de changement des conditions externes. Plus ce rapport sera faible, plus l'organisme se trouvera ťloignť de l'ťtat prťdit par les courbes de rťponse qui explicitent le taux de croissance mesurť ŗ l'ťtat stable en fonction du paramŤtre extťrieur (ces relations qui sont prťcisťment utilisťes dans les modŤles mathťmatiques). L'ťtude des transitoires qui accompagnent les changements d'ťtat (par exemple le rapport q) est donc essentielle. Celle-ci n'est possible que si l'on est capable d'une part d'imposer aux cellules un environnement continuellement variable de nutriment et de lumiŤre, et d'autre part d'en mesurer les diverses propriťtťs ŗ haute frťquence. Le forÁage qui conditionne en premier lieu la physiologie algale est le cycle diurne, car il induit le cycle cellulaire (temps qui s'ťcoule entre la naissance et la division) au cours duquel les phases de fixation de carbone et de division sont sťquentielles. Ainsi, une cellule n'utilise-t-elle qu'une partie de son cycle pour synthťtiser la matiŤre organique des cellules filles (Vaulot et al. 1987) . La production journaliŤre est donc l'intťgrale sur 24 heures de la fixation de carbone instantanťe, qui est variable au cours du cycle lumineux, mais aussi au cours du cycle cellulaire. La ressource en nutriment intervient car elle peut modifier le rapport des vitesses de chacun de ces cycles. Plus ils seront dťphasťs, plus le bilan de production journalier sera faible. On pressent donc que l'observation ŗ haute frťquence est importante car elle permet d'accťder ŗ l'ťchelle des cycles qui conditionnent le bilan de production sur le long terme. Ce n'est pas la seule raison. Le mode lumineux exploitť par les cellules dans la colonne d'eau diffťrera d'autant plus de la simple photopťriode que le gradient vertical de lumiŤre explorť par celles-ci sera important. Ainsi, existe-t-il une influence directe des propriťtťs hydrodynamiques sur la fixation de carbone ŗ travers la plasticitť du cycle cellulaire. Du point de vue de la modťlisation, ceci est trŤs intťressant car processus hydrodynamiques et cycle cellulaire peuvent Ítre chacun identifiťs par des constantes de temps caractťristiques. Par exemple, une cellule de convection peut Ítre caractťrisťe par son temps de rťvolution moyen, de mÍme qu'une espŤce phytoplanctonique par son temps de gťnťration. L'inverse de ces temps identifie la frťquence propre de ces systŤmes en interaction. La frťquence propre de l'entitť biologique peut, ŗ travers les processus d'adaptation, s'ajuster plus ou moins bien ŗ celle de la physique. Afin d'apprťhender cette plasticitť, il est nťcessaire de reconstituer un environnement oý les conditions de croissance sont continuellement variables, suivant un mode dictť par un schťma hydrodynamique prťcis.
Cette question est essentielle dans la modťlisation de rťseau trophique qui, ŗ juste titre, recherche ŗ rťduire la dimension des systŤmes traitťs. Une autre faÁon de poser la question est de savoir si l'on peut faire l'ťconomie d'un reprťsentation dťtaillťe. Il n'y a pas de rťponse univoque, suivant que l'on parle du systŤme rťel ou du modŤle. Quand on considŤre la rťalitť biologique (ou du moins ce que l'on en connaÓt), la question de l'importance des microphťnomŤnes ne se pose pas, car de toute faÁon, ils existent. La question est plutŰt de savoir si une composante ťmergente mesurable du systŤme, comme la production par exemple, traduit plus l'effet moyen rťsultant de ces microphťnomŤnes, ou bien leurs effets cumulťs. Dans le second cas, on doit les considťrer comme dťterminants, et ce ŗ deux points de vue. Ils le sont sur le bilan de matiŤre, car une intťgrale se calcule d'autant mieux que l'on a de points sur l'espace d'intťgration. Ils sont ťgalement dťterminants pour la trajectoire du systŤme car celui-ci ťvolue incrťmentalement en fonction de son ťtat. Mais il est probable, qu'en raison de l'important forÁage physique auxquels sont soumis les rťseaux trophiques, lequel forÁage est quasiment dťpourvu de rťtroaction, les trajectoires prises par le systŤme ťchappent en partie ŗ l'action des microphťnomŤnes. Cela est une supposition qui reste ŗ vťrifier. La question se pose aussi par rapport au modŤle. Dans quelle mesure une reprťsentation simpliste biaise-t-elle l'estimation des flux? Autrement-dit, une simple courbe de rťponse (cause-effet) est-elle prťfťrable ou non ŗ un petit systŤme de 3 ou 4 variables? Il n'y a pas de rťponse simple, car cela dťpend du forÁage appliquť (ŗ la limite, en rťgime stationnaire, les deux donnent le mÍme rťsultat). Mais ce qui est sŻr, c'est que le meilleur moyen de le savoir est de comparer les deux approches, et pour cela concevoir des modŤles de processus sur la base d'expťriences contrŰlťes. |
||||||||
|
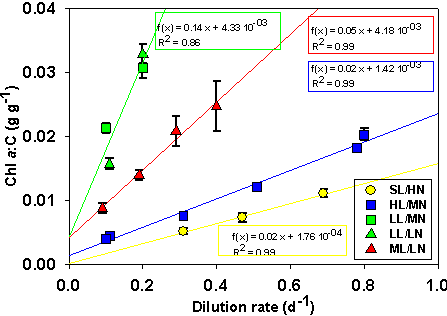
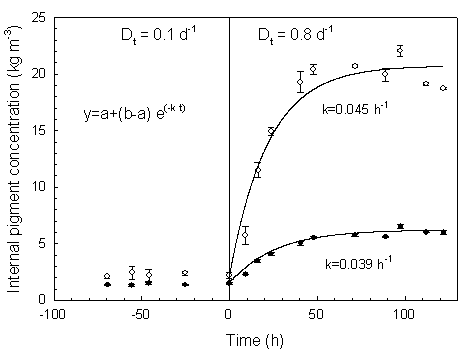
En mesurant les changements d'abondance d'ARN messagers, des protťines associťes aux complexes pigmentaires, et des pigments eux-mÍmes, il devient possible de dťterminer si les mťcanismes de photoadaptation identifiťs dans les algues contenant de la chlorophylle a et b et dans les chloroplastes des plantes supťrieures s'appliquent ťgalement ŗ d'autres groupes d'algues chez qui la composition pigmentaire et les antennes sont trŤs diffťrentes (c.a.d. les complexes phycobilisomes, fucoxanthine-chlorophylle, et pťridinine). En outre, il est nťcessaire d'examiner comment ce mťcanisme se produit lors de rťgimes transitoires de lumiŤre oý une sur-rťduction momentanťe de la CTE peut intervenir sur des pťriodes allant de la milliseconde ŗ la minute. Avec les outils de la biologie molťculaire, il est possible d'estimer les changements des pools d'ARNm, de protťines et de pigments qui rťsultent d'une modification de leur taux de synthŤse et de dťgradation, alors que l'acquisition en ligne de mesures biophysiques (fluorescence variable) permet d'estimer l'ťtat redox de la CTE photosynthťtique. Bien qu'il existe une information importante sur la physiologie de la photosynthŤse chez les algues et les cyanobactťries, celle-ci est actuellement de peu d'utilitť pour une validation rigoureuse des modŤles, notamment dans le cadre des conditions dynamiques existant dans le milieu naturel. A notre connaissance, la seule expťrience qui possŤde une information appropriťe permettant de tester un modŤle de la rťponse du rapport q ŗ une variation dynamique de lumiŤre a ťtť publiťe par Cullen et Lewis (1988), bien que cette expťrience ait ťtť conduite dans des conditions azotťes non limitantes. Encore faut-il mentionner que cette ťtude ne produit pas d'informations permettant de mettre en relation les rťponses photo-physiologiques et les mesures de fluorescence obtenues avec le PAM (Pulse Amplitude) ou le FRR (Fast Repetition Rate). De mÍme, la plupart des ťtudes conduites sur la rťgulation pigmentaire induite par un shift lumineux souffrent d'un manque critique de donnťes comme la concentration interne de carbone organique.
|
||||||||
|
Ces objectifs,
prťsentťs sťquentiellement pour des raisons
de clartť, sont nťanmoins interdťpendants. Ils visent ŗ rationaliser des actions
expťrimentales de laboratoire et de modťlisation de processus biologiques dans
la perspective de rťsoudre certains problŤmes inhťrents ŗ la
modťlisation des systŤmes complexes (rťseau trophique), dont le transfert
d'ťchelle n'est pas le moindre.
Il est nťcessaire d'ťtablir en amont de toute action une concertation entre physiciens, modťlisateurs et expťrimentateurs afin de dťfinir les protocoles d'expťriences au laboratoire les plus aptes ŗ fournir des rťsultats utiles ŗ la comprťhension et ŗ la modťlisation de la photoadaptation, processus central dans la production primaire, et sous reprťsentť dans les modŤles d'ťcosystŤme. Le problŤme est de dťfinir les modes de forÁage de lumiŤre, nutriment et ťventuellement de tempťrature ŗ spťcifier en entrťe aux systŤmes de culture pour que les cellules phytoplanctoniques mettent en œuvre les mťcanismes d'adaptation qu'elles utilisent effectivement lorsqu'elles ťvoluent dans une structure hydrodynamique bien dťfinie. Les diffťrentes situations hydrodynamiques identifiťes en Atlantique Nord lors de la campagne POMME constituent un cadre idťal pour cette ťtude.
Il s'agit d'approfondir la connaissance phťnomťnologique du processus, et d'apprťcier ses relations avec la croissance. Les effets synergiques de conditions variables de lumiŤre et de nutriment sur la productivitť photosynthťtique seront ťtudiťs ŗ l'aide de mťthodes biophysique et de biologie molťculaire avancťes. Ces mťthodes sont adaptťes pour identifier les mťcanismes impliquťs dans la rťgulation de la photosynthŤse au sein d'un milieu fluctuant.
∑
Mťthodes biophysiques et physiologiques. Les techniques de fluorescence
variable de la chlorophylle seront utilisťes pour quantifier les processus
photo-physiologiques, en dťterminant l'ťtat photochimique opťrationnel
du PhotosystŤme II, la dynamique de la capacitť photosynthťtique des
antennes (plus prťcisťment des LHS: light harvesting system), et la
cinťtique du transport d'ťlectrons entre les complexes photosynthťtiques
et la membranes thylakoÔde. Gr‚ce ŗ l'usage de fibres optiques, ces
mťthodes pourront Ítre utilisťes soit en ligne directement dans les
chťmostats soumis ŗ des conditions de croissance
variable, soit pour des mesures effectuťes ultťrieurement sur des ťchantillons
en cuves optiques. Les paramŤtres dťrivťs de la fluorescence variable
seront exploitťs pour la dťtermination de l'ťtat redox du pool de plastoquinone,
le dťtecteur supposť dťclencher la rťponse photoadaptative. Les bancs
de dťtection sensible seront constituťs par le Xe-PAM et le FRRF. ∑ Outils de biologie molťculaire et biochimique. L'expression des gŤnes photosynthťtiques en conditions de croissance variables sera suivie en quantifiant le niveau d'ARNm pour les gŤnes concernťs: psbA pour le centre rťactionnel du PhotosystŤme II, les gŤnes pour le principal complexe du LHS (lhc, pcbA ou fcp, en fonction de l'espŤce ťtudiťe), et rbcL pour l'expression d'une des grande sous-unitťs de la RUBISCO. Nous n'entrerons pas ŗ ce niveau dans le dťtail des procťdures d'ťlectrophorŤse, d'hybridation, et d'immuno-test.
Il s'agit plus prťcisťment des modŤles actuellement dťveloppťs ŗ Villefranche-sur-mer en collaboration avec l'INRIA ŗ travers le projet commun COMORE (ContrŰle et Modťlisation des Ressources Renouvelables). Ces modŤles sont conÁus pour rendre compte du comportement du rapport q et de la production dans un environnement co-limitť (nitrate-lumiŤre) et variable. Ce travail doit Ítre abordť sous trois aspects: 1. L'acquis phťnomťnologique explicitť dans le prťcťdent paragraphe doit servir ŗ dťfinir certains termes de ces modŤles. Il s'agira par exemple de prťciser si la dťgradation de la chlorophylle est uniquement un processus participant ŗ son renouvellement (dťgradation passive par dilution associťe ŗ la division cellulaire), ou s'il peut Ítre amplifiť dans des conditions de stress nutritif ou lumineux (dťgradation protťolytique). On recherchera ŗ identifier dans l'ensemble des mťcanismes impliquťs ceux qui sont essentiels. Il s'agit bien ťvidemment de ne pas utiliser toute l'information extraite des expťriences pour alimenter des modŤles complexes surdimensionnťs. Au contraire on recherchera d'une part ŗ dťfinir les critŤres permettant d'effectuer des choix de simplification dans les variables pertinentes ŗ retenir, et d'autre part ŗ prťciser les limites d'utilisation du modŤle sur la base des mťcanismes retenus ou abandonnťs. 2. Il n'y a qu'en connaissant bien la complexitť du systŤme rťel qu'on peut en trouver une image thťorique (modŤle) simplifiťe. L'effort portera donc sur la recherche de la structure mathťmatique la plus simple (systŤme de faible dimension avec peu de paramŤtres) qui rende compte des propriťtťs jugťes indispensables pour une reprťsentativitť minimale acceptable. La dťfinition de ce qui est acceptable ou non s'accorde sur la connaissance acquise sur le systŤme rťel, c'est-ŗ-dire sur les donnťes de campagne, aussi bien physiques que biologiques. 3. La validation des modŤles doit accompagner leur conception et non la suivre. Travailler la paramťtrisation pour ajuster le modŤle aux observations ne constitue pas une validation suffisante. Pour qu'un modŤle soit structurellement validť, il est nťcessaire qu'aucune des trajectoires qu'il peut produire ne sorte de ce qui est potentiellement observable sur le systŤme rťel. La proposition symťtrique est qu'un systŤme biologique ne peut pas occuper tout l'espace des ťtats, mais que ses propriťtťs intrinsŤques le contraignent ŗ ťvoluer au sein d'un domaine spťcifique de cet espace. L'ťtude mathťmatique des systŤmes d'ťquations diffťrentielles permet de dťfinir ce domaine, qui, pour une grande part, est indiffťrent ŗ la formulation mathťmatique des ťquations et ŗ leur paramťtrisation. Une ťtude structurelle du modŤle permet, en amont de toute simulation, d'orienter sa dťfinition pour que ses propriťtťs qualitatives satisfassent celles du systŤme biologique. Les recherches expťrimentales menťes en chemostat se prÍtent tout ŗ fait ŗ ce genre de technique, car les termes d'entrťe sortie (il s'agit d'un systŤme ouvert), que l'on peut assimiler ŗ des termes physiques, confŤrent ŗ l'ensemble (systŤme biologique dans le chemostat) des propriťtťs inhťrentes particuliŤres que les modŤles doivent satisfaire impťrativement. I.4.4. Retour au système réel: couplage physique-biologie Le dťveloppement de modŤles de processus ne peut s'arrÍter au stade de leur validation expťrimentale, qui n'offre qu'un cadre restreint ŗ leur application future. L'ťtape ultime avant leur insertion dans des structures importantes (modŤles d'ťcosystŤme) nťcessite un test de faisabilitť et d'utilitť. Le test de faisabilitť doit rťsoudre tous les problŤmes que l'on rencontre lors du couplage d'un modŤle hydrodynamique avec un modŤle physique. Il est prťfťrable d'effectuer cette opťration en l'absence de toute autre variable qui ne participe pas directement au processus impliquť. Pour ťtudier le comportement d'un modŤle de photoadaptation au sein d'une structure physique, il n'est pas nťcessaire d'y inclure ni les bactťries, ni le zooplancton, car il ne s'agit bien ťvidemment pas, ŗ ce stade, de reproduire le systŤme rťel. On se sert de la structure physique comme d'un chťmostat: pour expťrimenter sur une partie du systŤme, ce qui simplifie grandement l'approche de validation. Le test d'utilitť doit permettre de dťterminer deux choses. PremiŤrement le gain que procure un modŤle de photoadaptation par rapport ŗ des courbes de rťponse associťes par des lois empiriques (minimum, multiplicatif, etc.): le comportement oý les bilans produits par ce modŤle sont-ils radicalement diffťrents du systŤme "courbe de rťponse"? La rťponse n'ťtant probablement pas simple, il s'agira de dťfinir la classe de modŤle ŗ utiliser en fonction du forÁage hydrodynamique en vigueur. Dans tous les cas, il est importance d'estimer le biais auquel on s'expose en se contentant soit d'une reprťsentation trop simplifiťe, soit au contraire d'un modŤle sur-paramťtrť. |
||||||||
† Bernard O (1995) Etude expťrimentale et thťorique de la croissance de Dunaliella tertiolecta (chlorophyceae) soumise ŗ une limitation variable de nitrate: utilisation de la dynamique transitoire pour la conception et la validation des modŤles. P.M. Curie, Paris VI, pp 200 Bernard O, Gouzť JL (1995) Transient behavior of biological loop models with application to the Droop model. Math. Biosciences 127:19-43 Bernard O, Gouzť JL (1996) Transient behavior of biological models as a tool of qualitative validation - Application to the Droop model and to a N-P-Z model. J. Biol. Syst. 4:303-314 Bernard O, Malara G, Sciandra A (1996) The effects of a controlled fluctuating nutrient environment on continuous cultures of phytoplankton monitored by a computer. J. Exp. Mar. Biol. Ecol 197:263-278 Bruyant F, Babin M, Sciandra A, Marie D, Genty B, Claustre H, Blanchot J, Bricaud A, Rippka R, Boulben S, Louis F, Partensky F (2001) An axenic cyclostat of Prochlorococcus PCC 9511 with a simulator of natural light regimes. J. Appl. Phycol. 13:135-142 Collos Y, Sciandra A, Martin-Jťzťquel V, Slawick G, Garcia N (1998) Dilution of 15N in Dunaliella tertiolecta by uptake of an unidentified nitrogen compound following nitrate exhaustion. C. R. Acad. Sci. 321:673-677 Cullen JJ, Geider RJ, Ishizaka J, Kiefer
DA, Marra J, Sakshaug E, Raven JA (1993) Toward a general description
of phytoplankton growth for biogeochemical models. In: Evans GT,
Fasham MRJ (eds) Toward a model of ocean biogeochemical processes. Springer-Verlag, pp 153-176 Cullen JJ, Lewis MR (1988) The kinetics of algal photoadaptation in the context of vertical mixing. J. Plankton Res. 10:1039-1063 Davidson K, Wood G, John EH, Flynn KJ (1999) An investigation of non-steady-state algal growth. I. An experimental model ecosystem. J. Plankton Res. 21:811-837 Escoubas, J.M., Lomas, M., La Roche, J. et Falkowski, P.G. (1995) Light intensity regulation of cab gene transcription is signaled by the redox state of the plastoquinone pool. Proc. Natl. Acad. Sci. 92:10237-41 Eveillard
D (2000) Modťlisation de l'effet de la limitation conjugťe de la lumiŤre et
de l'azote sur la croissance autotrophe. Universitť P.M. Curie, Paris,
pp 32 Flynn KJ, Marshall H, Geider RJ (2001) A comparison of two N-irradiance interaction models of phytoplankton growth. Limnol. Oceanogr. 46:1794-1802 Fujita, Y., A. Murakami, et al. (1994). The Molecular Biology of Cyanobacteria, Kluwer, Dordrecht. Geider RJ (1987) Light and temperature dependence of the carbon to chlorophyll a ratio in microalgae and cyanobacteria: Implications for physiology and growth of phytoplankton. New Phytol. 106:1-34 Geider RJ, MacIntyre HL, Kana TM (1997) Dynamic model of phytoplankton growth and acclimation: responses of the balanced growth rate and the chlorophyll a : carbon ratio to light, nutrient-limitation and temperature. Mar. Ecol. Prog. Ser. 148:187-200 Geider RJ, MacIntyre HL, Kana TM (1998) A dynamic regulatory model of phytoplanktonic acclimation to light, nutrients, and temperature. Limnol. Oceanogr. 43:679-694 Kim, J. and S. P. Mayfield (1997). Protein
disulfide isomerase as a regulator of chloroplast translational activation. Science 278: 1954-1957. Kolber, Z. S., O. Prasil et Falkowski, P. (1998). Measurements of variable chlorophyll fluorescence using fast repetition rate techniques: defining methodology and experimental protocols. Biochimica et Biophysica Acta 1367(1-3): 88-106. Lande R, Lewis MR (1989) Models of photoadaptation and photosynthesis by algal cells in a turbulent mixed layer. Deep-Sea Res. (a Oceanogr. Res. Pap.). 36:1161-1175 Le Floc'h E, Malara G, Sciandra A (in revision) An automatic device for in vivo absorption spectra acquisition in phytoplanktonic cultures: application to the study of photoadaptation to light and nutrient variations. J. Applied Phycol. Le Floc'h E, Sciandra A (2000) Adaptation of pigment composition to interactive effects of light and nitrogen in Rhodomonas salina (Cryptophyceae) ASLO meeting, Abulquerque Lewis MR, Horne EPW, Cullen JJ, Oakey NS, Platt T (1984) Turbulent motions may control phytoplankton photosynthesis in the upper ocean. Nature 311:49-50 Malara G, Sciandra A (1991) A multiparameter phytoplanktonic culture system driven by microcomputer. J. Applied Phycol. 3:235-241 Marra J (1978a) Effect of short-term variations in light intensity on photosynthesis of marine phytoplankter: a laboratory simulation study. Mar. Biol. 46:191-202 Marra J (1978b) Phytoplankton photosynthetic
response to vertical movement in a mixed layer. Mar. Biol. 46:203-208 Pawlowski
L (2001) Modťlisation des effets conjuguťs azote/lumiŤre sur la croissance
phytoplanctonique. Universitť P.M. Curie, Paris, pp 31 Pawlowski L, Bernard O, Le Floc'h E, Sciandra A (2002) Qualitative behaviour of phytoplankton growth model in photobioreactor 15th Word Congress, Barcelona, Spain Peperzak, L., Colijn, F., Gieskes, W.W.C. et Peeters, J.C.H. (1998) Development of the diatom-Phaeocystis spring bloom in the Dutch coastal zone on the North Sea. J. Plankton Res. 20: 517-537 Schwarz, R. and A. A. Grossman (1998). A response regulator of cyanobacteria integrates diverse environmental signals and is critical for survival under extreme conditions. Proceedings of the National Academy of Science USA 95: 11008-11013. Sciandra A (1991) Coupling and uncoupling between nitrate uptake and growth rate in Prorocentrum minimum (Dinophyceae) under different frequencies of pulsed nitrate supply. Mar. Ecol. Prog. Ser. 72:261-269 Sciandra A, Amara R (1994) Effects of nitrogen limitation on growth and nitrite excretion rates of the dinoflagellate Prorocentrum minimum. Mar. Ecol. Prog. Ser. 105:301-309 Sciandra A, Gostan J, Collos Y, Descolas-Gros C, Leboulanger C, Martin-Jťzťquel V, Denis M, LefŤvre D, Copin-Montťgut C, Avril B (1997) Growth compensating phenomena in continuous cultures of Dunaliella tertiolecta limited simultaneously by light and nitrate. Limnol. Oceanogr. 42:1325-1339 Sciandra A, Lazzara L, Claustre H, Babin M (2000) Responses of the growth rate, pigment composition and optical properties of Cryptomonas sp. to light and nitrogen stresses. Mar. Ecol. Prog. Ser. 201:107-120 Sciandra A, Ramani P (1994) The steady states of continuous cultures with low rates of medium renewal per cell. J. Exp. Mar. Biol. Ecol. 178:1-15 Stramski D, Sciandra A, Claustre H (2002) Variations in the optical properties of the marine diatom Thalassiosira pseudonana induced by various sources of growth limitation. Limnol. Oceanogr. 47:00-00 Vaulot D, R.J. O, Merkel S, Chisholm SW (1987) Cell-cycle response to nutrient starvation in two phytoplankton species, Thalassiosira weissflogii and Hymenomonas carterae. Mar. Biol. 95:625-630 Zakardjian, B. and Prieur, L. 1998. Biological and chemical signs of upward motions in permanent geostrophic fronts of the Western Mediterranean. J. Geophys. Res., 103, C12, 27849 - 27866. Zonneveld C (1998) A cell-based model
for the chlorophyll a to carbon ratio in phytoplankton. Ecol. Model. 113:55-70. |
||||||||