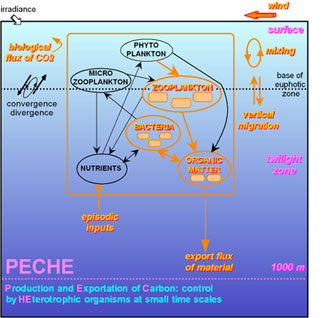
Production and Exportation of Carbon : control by HEterotrophic organisms at small time scales
Valérie
ANDERSEN |
Madelein GOUTX |
The
aim of this project is then to examine short-term temporal variability
of
the primary
production and export
fluxes to depth. It is focussed on the study of the control by heterotrophic
organisms (from bacteria to macrozooplankton) and of the response
of the system to episodic events during a seasonal transition period,
along the vertical dimension. ††††††††††† This
project is essentially based on in
situ observations which will be carried out in different seasons:
two cruises, in April (period of abundant zooplankton biomass) and
July (period of minimum downward flux), and a one month time-series
in autumn (September-October). This project also includes some modelling
studies (1-D coupled physical-biological models). ††††††††††† The
specific objectives of the three observation periods are: (1) Characterisation of the relative impact
of zooplankton and bacteria (and their associated ecto-enzymes) in
the mineralization of the organic matter, (2) Quantification of the variability
of mineralisation† with the
structure of the heterotrophic web and the depth, (3) Estimation of the response of the
ecosystem to transient episodes, such as wind events, at small time
scales of a few hours to a few days during the seasonal transition
from summer to autumn; this aspect will be studied during a multidisciplinary
cruise which will integrate the process studies performed at the other
seasons. ††††††††††† Our
overall† goal is to improve
the budget of carbon production and exportation throughout the water
column (from the surface to 1000 m depth) and to determine the relative
contribution of the different heterotrophic processes and of the physical
forcings at small time scales. ††††††††††† This project will also provide a multidisciplinary data set which will be used to parameterize, constrain and validate a variety of physical-biological models |
Le projet PECHE vise ŗ une meilleure apprťciation du contrŰle de la production primaire et du flux exportť en profondeur. Il est axť sur líťtude du contrŰle par les organismes hťtťrotrophes (de la bactťrie au macrozooplancton) et des rťponses du systŤme aux perturbations impulsionnelles. Ce projet est basť principalement sur des observations in situ, mais fait aussi appel ŗ des
travaux de modťlisation 1-D couplťe physique-biologie. ††††††††††† A partir de ce bilan des
connaissances sur le contrŰle des flux de matiŤre par les organismes
hťtťrotrophes et le rŰle de la petite ťchelle temporelle, les principaux
objectifs visťs par le projet PECHE peuvent Ítre exprimťs de la faÁon suivante
: †- Quel est le rŰle du broutage et de
l'excrťtion du zooplancton dans le contrŰle de la production primaire et dans
la mise ŗ disposition de ressources organiques pour le compartiment bactťrien
? - Comment
la diversitť structurelle et fonctionnelle du rťseau hťtťrotrophe
influence-tíelle le flux exportť en profondeur ? - Quelle
est la rťponse de l'ťcosystŤme ŗ l'impact de perturbations impulsionnelles telles
que les coups de vent, rťponse non seulement ŗ l'ťchelle de quelques heures ŗ
quelques jours mais aussi pendant la transition saisonniŤre ? ††††††††††† L'objectif final est d'amťliorer le bilan de la production et de l'exportation de carbone sur la colonne d'eau de la surface jusqu'en profondeur (1000 m), et de dťterminer la part respective des diffťrents processus hťtťrotrophes et des forÁages physiques ŗ petite ťchelle de temps. |
|
|
|
|
|
The project PECHE is designed to examine
the natural variability of the structure and the dynamics of the pelagic
ecosystems at small time scales and in response to transient events.
The project adresses a major question concerning the functioning of
biological systems in the open ocean and the biogeochemical responses
to global change. ††††††††††† In
the pelagic ecosystem, bottom-up control of phytoplankton dynamics
has long been considered as the most important factor in regulating
primary production. The Top-down control, that is predation by herbivores
as well as by carnivores and the pivotal role of zooplankton grazing
on phytoplankton composition dynamics has been only recently emphasized.
While it is clear that the functioning of the trophic web depends then
on the equilibrium between bottom-up and top-down controls, the relative
importance of these controls is a matter of debate. ††††††††††† Moreover,
the trophic web presents a large structural and functional diversity,
which determines the efficiency of the biological carbon pumping. Schematically,
the trophic web can be divided into two main pathways : (i) the microbial
loop with controls the circulation and retention of matter in the euphotic
zone (e.g. bacterial mineralization of particulate and dissolved organic
matter, predation and excretion of ciliates) and (ii) the 'macrobial'
web which plays a determinant role in the export flux (e.g. production
of large fecal pellets with high sedimentation rates, diel vertical
migration). ††††††††††† The microbial loop is commonly recognized as a major pathway in carbon transfer and regulation in biological systems. At its base, bacteria assimilate and re-mineralize detrital and dissolved organic matter. The importance of chemical composition of organic matter on bacterial metabolisms and dynamics is well known. However, organic matter in the ocean is mainly under the form of polymeric substrates and ecto-enzymatic hydrolysis by specific enzymes is a preliminary step to assimilation of most substrates.Thus, relationships between diversity and functionality within bacterial community is a major challenge for studying the impact of bacterial processes on fluxes of matter and energy.
††††††††††† †††††††† Biological structure strongly coupled with physical structures varies over a large range of time and space scales in the ocean. Importance of the processes occurring at small time scales have been progressively emphasized. In fact, the response of the system to transient episodes are often complex and non-linear. Reliable budgets for the long term periods (seasons to years) cannot be calculated if data are lacking on the short-term variances and covariances. Seasonal transition periods, characteristic of temperate areas, appear particularly critical, as they may influence both seasonal and interannual variability. They are themselves controlled by interactive and successive rapid processes, from hours to a few days. |
|
|
|
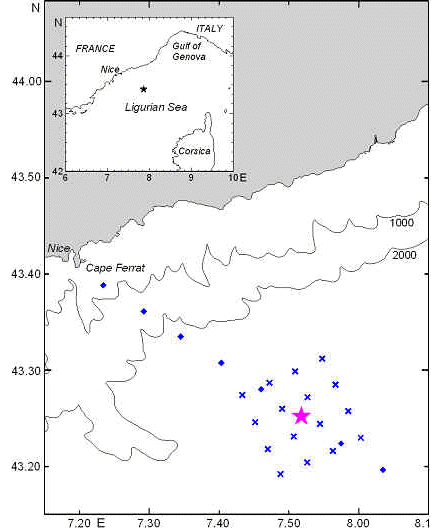
| This study, based on vertical processes,
will be conducted in the central zone of the Ligurian Sea (NW Mediterranean)
where advective movements are negligible, in the vicinity of the long-term
time-series station DYFAMED. The multidisciplinary results of this
study, experimental as well as theoretical, could then be extrapolated
to other ocean areas, temperate ones particularly. - Mineralisation of organic matter. Cruises PROPECHE
(April and July 2003) ††††††††††† Mineralisation
of organic matter depends on stratification, nutrient conditions and
associated trophic conditions. Zooplankton community outputs can significantly
modify the impact of environmental conditions on the mineralisation
process. Thus, interactions between zooplankton and bacteria† will
be† specifically examined during
the PROPECHE cruises (PROcess
of organic matter mineralisation within PECHE), using a pluridisciplinary
approach. Ė Along vertical profiles (0-1000 m),
we will examine the distribution of chemical (nutrients, CHN, DOC,
DON), biochemical (OM composition of proteins, carbohydrates, lipids,
fulvic acids) and microbiological (activities and diversity) parameters
in the dissolved and suspended matter collected during day and night
sampling. - Aggregates will be collected using a
special detritus sampler; fresh faecal pellets will be obtained from
incubation of different sized zooplankton groups sampled with nets. - Chemical, biochemical and microbiological
characteristics of these particles and aggregates will be followed
during short incubation times (< 36 h). - Bacterial biomass, bacterial production,
and bacterial community structure (Finger Printing, CE-SSCP) will be
also examined. - The chemical analysis of biopolymers
will be coupled with the analysis of associated monomers and to the
estimation of hydrolysis rates (using fluorescent analogs and polymeric
substrates). ††††††††††† This
approach will be also conducted during the DYNAPROC 2 cruise. Thus,
these observations at three periods characterized by different trophic
organisations would provide insight on zooplankton/bacteria interactions
and basic parameters for mineralization process modelling.†
- Small time-scales response of the ecosystem to episodic
events during the transition from summer to autumn. Cruise DYNAPROC
2 (September-October 2004)
††††††††††† Wind-induced
mixing events, such as those occurring in autumn in NW Mediterranean,
can increase availability of nitrates in the euphotic layer. What is
the influence of this enhancement on the autumnal bloom of phytoplankton
? On the other hand, microphageous organisms (such as pteropod molluscs
and pyrosomids) occur in large densities at this period and could control
the phytoplankton community which, in autumn, consists mainly of small-sized
cells. ††††††††††† Answers
to these questions will be specifically addressed during the cruise
DYNAPROC 2 (DYNAmics of rapid PROCessses in the water column). ††††††††††† The
main observations will be done near a fixed station and monitor: daily
variation of phytoplankton biomass and related physical and chemical
parameters, and changes in the trophic web structure and functioning
(e.g. phytoplankton, microbial loop, large filter-feeders). Most of
these biogeochemical studies will be based on measurements at short
time scales (4 to 12 hours) during 48-hours cycles. In
situ observations and sampling will be performed with various technics,
such as CTD and 24-bottle rosette, multiple and opening closing net,
underwater video profiler, moored system for biological fluxes of CO2.
The processes related to primary production will be explored in the
0-200 m water column, those depending on heterotrophic activities from
the surface to 1000 m depth. These process studies will concern: primary
production, physiological rates of zooplankton ††††††††††† To
determine the hydrological environment of the fixed station, a grid
of 16 stations, centred on the fixed station, will be occupied at least
four times during the cruise. ††††††††††† In
order to document the seasonal transition period and to catch the effects
of the wind event, which can be delayed in the zooplankton response
compared to the phytoplankton response, the cruise will be planned
to last for at least 4 weeks. ††††††††††† This cruise will complement the first DYNAPROC cruise (May 1995) which documented the effects of episodic wind events on the transition from the mesotrophic system to an oligotrophic one. A 1-D physical-biological model, first applied to the DYNAPROC data and including vertical advection, will be applied to the time series of DYNAPROC 2 and tested, before to being run at a longer larger time scale. |
|
Les recherches
rťcentes et les projets en ocťanographie biologique síarticulent autour
de plusieurs thŤmes majeurs dont : (1) les rťponses des systŤmes biologiques
aux variations globales, en particulier climatiques, et (2) les interactions
entre les cycles biogťochimiques et la structure et le fonctionnement
des ťcosystŤmes. ††††††††††† Ce contexte soulŤve des questions
majeures sur la variabilitť naturelle
de la structure et de la dynamique de líťcosystŤme pťlagique, questions
auxquelles notre projet vise ŗ apporter des ťlťments de rťponse.
|
|
††† Les images satellitaires
montrent que la physique de líocťan joue un rŰle essentiel sur la distribution
spatiale et temporelle de la biomasse phytoplanctonique. Ainsi, les
foyers de production ťlevťe correspondent ŗ des zones díenrichissement
de la couche euphotique en sels nutritifs. Ce contrŰle par les ressources
(bottom-up control des anglo-saxons) a longtemps ťtť considťrť comme
dťterminant pour la dynamique du phytoplancton, et nettement plus important
que le contrŰle par le niveau trophique supťrieur ou prťdation (top-down
control). ††††††††††† Líimportance
du contrŰle par la prťdation, tant des herbivores que des carnivores,
et le rŰle pivot du broutage du zooplancton sur la dynamique du phytoplancton
níont ťtť soulignťs que depuis quelques annťes, pour líocťan ouvert
en particulier (e.g. Banse, 1994, 1995; Verity & Smetacek, 1996).
Líactivitť de ces consommateurs secondaires aurait des consťquences
importantes dans la rťgulation du flux de carbone circulant ŗ travers
le rťseau microbien (Christaki & Van Wambeke, 1995). ††††††††††† Ainsi le niveau de fermeture (prťdation)
dans les modŤles de rťseau trophique conditionne le comportement de
toutes les variables. Par exemple, Steele & Henderson (1992) ont
rappelť líimportance de la formulation de ce terme au niveau des herbivores
et Frost & Franzen (1992) ont montrť que leur modŤle ŗ líťtat stationnaire
ne reproduisait les caractťristiques de la dynamique du phytoplancton
observťes in situ que lorsque deux niveaux de carnivores ťtaient
considťrťs. ††††††††††† Le fonctionnement
du rťseau trophique dťpend donc de líťquilibre entre le contrŰle par
les ressources et le contrŰle par la prťdation; líimportance relative
de ces contrŰles est encore une question dťbattue (par exemple Longhurst,
1991; Banse, 1995; Franks, 2001; Marine Zooplankton Colloquium 2, 2001). ††††††††††† La quantification
du contrŰle de la dynamique de líťcosystŤme par la prťdation, tant
par les herbivores que par les carnivores, apparaÓt centrale si líon
veut interprťter les observations globales et intťgrťes et prťdire
les biomasses phytoplanctoniques et leurs variations ŗ diffťrentes ťchelles
spatiales et temporelles. |
|
Au sein
du rťseau trophique, on peut distinguer
schťmatiquement deux voies principales : la voie microbienne (pico-
et nanophytoplancton, bactťries, ciliťs) et la voie macrobienne composťe
díorganismes de grande taille (diatomťes, copťpodes, salpes,...). Líinfluence
principale de ces deux voies sur la pompe biologique du CO2 peut Ítre
rťsumťe en síappuyant sur Longhurst (1991) qui a divisť schťmatiquement
la pompe biologique en trois processus ťcologiques : (1) une pompe
rotative (rotary pump) qui correspond ŗ la circulation et ŗ la rťtention
de la matiŤre dans la zone euphotique sous líaction du rťseau microbien,
(2) une pompe díArchimŤde (Archimedian pump) qui correspond au flux
vertical de particules (pelotes fťcales, agrťgats, coquilles calcifiťes,...)
produites par le rťseau macrobien, (3) une pompe alternative (reciprocating
pump) reprťsentťe par les organismes migrateurs.
††††††††††† Le
rťseau microbien est ubiquiste en milieu marin et les nombreuses ťtudes
qui lui ont ťtť consacrťes ces vingt derniŤres annťes ont considťrablement
augmentť notre connaissance des flux de matiŤre et díťnergie dans
les† couches superficielles (Azam, 1998). ††††††††††† Les bactťries jouent un rŰle essentiel
dans le rťseau trophique par leur capacitť ŗ assimiler et reminťraliser
la matiŤre organique dťtritique et/ou dissoute (MO). Ce sont en effet
les rares organismes capables de rťcupťrer des concentrations extrÍmement
faibles de molťcules organiques dispersťes dans le milieu sous forme
dissoute et/ou colloÔdale. La respiration d'une partie de ces substrats
en CO2 s'accompagne de la remise ŗ disposition des sels
minťraux ŗ l'ensemble du rťseau trophique. La fraction assimilťe produit
une biomasse bactťrienne ŗ la base du rťseau trophique; cette biomasse
est utilisťe par le nano- et le microzooplancton qui sont ŗ leur tour
la proie du zooplancton macrobien.† Une fraction de la MO qui, sans l'intervention
bactťrienne serait perdue pour le systŤme, est ainsi rťinjectťe dans
le rťseau trophique. Si de nombreux travaux montrent que la concentration
et la composition ťlťmentaire des substrats sont des paramŤtres importants
dans la rťgulation de la dynamique bactťrienne (cf. Touratier et al.,
2001), les substrats considťrťs sont le plus souvent de petites molťcules
organiques. ††††††††††† Ainsi, la relation MO-bactťries est
encore dťfinie de faÁon peu prťcise. Une grande partie de la MO, qu'il
s'agisse de dťtritus planctoniques, de pelotes fťcales ou d'exsudats
frais, se trouve dans le milieu sous forme de biopolymŤres de grande
taille. En effet, seules les molťcules de faible poids molťculaire
(<600) sont directement accessibles aux bactťries. C'est l'hydrolyse
par les ecto-enzymes bactťriennes qui rťalise le clivage des biopolymŤres
en monomŤres assimilables. Allant ŗ l'encontre de cette mise ŗ disposition
des polymŤres et des monomŤres, diffťrents processus, tels que condensations
chimiques, protection physique par une fraction minťrale ou agrťgations
biologiques, par le zooplancton notamment, entraÓnent la nťoformation
de molťcules plus ou moins accessibles aux enzymes bactťriennes, ŗ partir
de la MO biogŤne (Carlson et al., 1985; Amon & Benner, 1994;† Amstrong
et al., 2002). ††††††††††† Líefficacitť de minťralisation du
rťseau microbien ne peut donc Ítre comprise et estimťe qu'ŗ partir
díune approche pluridisciplinaire tenant compte ŗ la fois de la variabilitť des
caractťristiques chimiques de la matiŤre organique et de la diversitť des
espŤces bactťriennes et de leurs fonctionnalitťs. Cette relation diversitť/fonctionnalitť des
communautťs bactťriennes est aujourdíhui un enjeu majeur pour l'ťtude
de líimpact des processus bactťriens dans les flux de matiŤre et d'ťnergie
(Cottrell & Kirchman, 2000a). Enfin, la minťralisation s'exerce ŗ tous
les niveaux de la colonne d'eau, depuis la surface jusqu'en profondeur.
La spťcificitť des milieux mťso- et bathypťlagique doit donc Ítre intťgrťe ŗ l'ťtude
de ce processus.
††††††††††† Cette "pompe
rotative", gouvernťe par le rťseau microbien, est ťtroitement
liťe ŗ la ďpompe díArchimŤdeĒ et ŗ la ďpompe alternativeĒ dont la signification
potentielle nťcessite une meilleure comprťhension de la structure et du fonctionnement du rťseau macrobien.
††††††††††† Le zooplancton
macrobien prťsente une grande diversitť structurelle et fonctionnelle.
Le rapport de taille entre prťdateur et proie montre une trŤs grande
variabilitť chez le zooplancton macrobien (Longhurst, 1991; Fortier
et al., 1994). Par exemple, certains organismes, tels que les crustacťs
(copťpodes, euphausiacťs) sont capables de se nourrir sur les grandes
cellules phytoplanctoniques (diatomťes) qui ne sont pas accessibles
au microzooplancton. Par contre, certains groupes díorganismes gťlatineux,
tels que salpes, appendiculaires et mollusques ptťropodes, exploitent
efficacement le nanophytoplancton et sont donc en compťtition avec
le microzooplancton. ††††††††††† Díautre part,
les niveaux trophiques ne se limitent pas aux herbivores et carnivores
stricts, mais comprennent aussi des omnivores et des dťtritivores,
díoý líabandon du concept de chaÓne trophique au profit de celui de
rťseau trophique. Si la diversitť du zooplancton macrobien est reconnue,
la structure de líťcosystŤme et la succession des espŤces dominantes
sont encore peu prises en compte dans la prťdiction des flux de carbone. ††††††††††† Le zooplancton
macrobien est aussi ŗ líorigine de la majoritť des grandes particules
exportťes en profondeur. Ce flux contrŰle le cycle des ťlťments chimiques
mais il fournit aussi la quasi-totalitť de l'apport ťnergťtique ŗ la
faune des grands fonds (e.g. Coale & Bruland, 1985; Fowler & Knauer,
1986). Ces particules, telles les pelotes fťcales des salpes, peuvent
prťsenter des vitesses de sťdimentation trŤs ťlevťes. Elles sont produites ŗ toutes
les profondeurs, avec en particulier une remobilisation du matťriel
dťtritique par diffťrents types díorganismes zooplanctoniques. ††††††††††† Une fraction
de la matiŤre produite en surface est ťgalement transportťe en profondeur
de maniŤre active par les migrateurs et cette exportation est assurťe
principalement par les organismes de grande taille. L'effet de cette
migration verticale nycthťmťrale du zooplancton sur la dynamique de
l'ťcosystŤme et les flux de matiŤre organique et inorganique est encore
mal quantifiť, bien que son importance soit largement reconnue (e.g.
Angel, 1989; Longhurst, 1991; Banse, 1995; Marine Zooplankton Colloquium
2, 2001). ††††††††††† La composition
spťcifique du rťseau trophique, ŗ tous ses ťchelons, est donc un facteur
dťterminant líefficacitť de la pompe biologique du CO2.
|
|
††††††††††† La plus grande
partie de la matiŤre organique synthťtisťe par les producteurs primaires
dans la couche euphotique est recyclťe dans la zone ťpipťlagique ŗ travers
les rťseaux trophiques dynamiques et seulement quelques % de cette
matiŤre est exportťe au-delŗ de 1000 m (Lampitt & Antia, 1997).
Or seule cette fraction de la matiŤre organique entraÓnťe vers les
eaux profondes soustrait du CO2 ŗ l'atmosphŤre ŗ long ou
moyen terme (Ittekott et al. 1996). ††††††††††† Les particules
les plus denses, dont la densitť leur permet franchir les gradients
physiques des eaux superficielles et de la colonne d'eau, constituent
la majeur partie du flux particulaire. Les organismes intacts peuvent
reprťsenter jusqu'ŗ 50% de ce flux qui comprend ťgalement des pelotes
fťcales d'origine zooplanctonique et des agrťgats de diffťrentes sources
et tailles. Ces agrťgats constituent des micro-environnements trŤs
rťactifs associant bactťries endogŤnes et MO. Ainsi les caractťristiques ŗ líťchelle
particulaire, en terme de MO et de diversitť des bactťries (auxquelles
vient síajouter la prťsence de nanoflagellťs et de ciliťs hťtťrotrophes)
peuvent dťterminer le rŰle spťcifique des bactťries et du zooplancton
dans les vitesses de sťdimentation et le contrŰle du flux particulaire.
Le flux particulaire constitue aussi un des rares apports de dťtritus
phytoplanctoniques au sťdiment profond. ††††††††††† D'autre part,
la convexion des masses d'eau entraÓne en profondeur la fraction du
carbone organique biogŤne non assimilťe dans le rťseau trophique et
accumulťe en surface sous forme dissoute. Une fraction de cette MO
rejoint le flux particulaire par adsorption et rť-agrťgation. Enfin,
une fraction perd sa biodisponibilitť par transformation en molťcules
plus rťfractaires. ††††††††††† Il est donc indispensable
de bien comprendre la dynamique du flux particulaire ŗ l'aide d'outils
nouveaux, pour mieux contraindre les flux biologiques actuels. La connaissance
des flux "modernes" et des outils de leur traÁabilitť peut
trouver des applications dans l'identification des processus biogťochimiques "anciens". ††††††††††† Les marqueurs
organiques ont par dťfinition la capacitť de conserver les signaux
des sources et transformations subies par la matiŤre organique biogŤne
jusquíŗ sa minťralisation ou sa prťservation. Cíest ce qui leur confŤre
la possibilitť de dťcrire et comprendre les flux majeurs díťchanges
entre les rťservoirs de líatmosphŤre, líocťan et le sťdiment (Wakeman & Lee,
1993). Au sein du rťseau trophique, ces marqueurs ont pour origine
les bio-polymŤres des cellules vivantes et des dťtritus. La mise ŗ disposition
des bio-polymŤres ŗ des systŤmes enzymatiques plus ou moins diversifiťs
du rťseau trophique va, comme nous líavons vu, prťluder ŗ líassimilation
et ŗ la respiration bactťriennes. Les processus de dissolution, re-condensation
ou bien agrťgation particulaire influencent largement le flux díexportation,
en accťlťrant la mise ŗ disposition des bio-polymŤres ou au contraire
en les protťgeant de líaction des ecto-enzymes (dont l'activitť est
par ailleurs mesurable).† ††††††††††† Ainsi l'ťtude
du comportement de marqueurs organiques doit inclure la mesure de la
vitesse rťelle de la production de leur mťtabolites de dťgradation
dans le milieu par les ecto-enzymes bactťriennes, de leur assimilation
par les bactťries et/ou de leur accumulation dans le milieu. Cette
approche constitue une dťmarche biogťochimique nouvelle, conciliant
les approches des microbiologistes et des gťochimistes. Elle peut Ítre
appliquťe ŗ diffťrents modŤles molťculaires tels que les lipides de
type glycťryl-esters, leurs mťtabolites (les acides gras), les activitťs
lipolytiques et les lipases. |
|
† Dans líocťan, les structures
physiques et les structures biologiques varient dans une large gamme
díťchelles temporelles et spatiales (e.g. Haury et al., 1978). Il est
fondamental de comprendre le couplage entre les ťchelles, des plus
petites aux plus grandes, et díen quantifier les consťquences. Au cours
des quinze derniŤres annťes, les ocťanographes ont progressivement
pris conscience de líimportance des processus se dťroulant aux petites ťchelles
de temps (Marine Zooplankton Colloquium 1, 1989; Platt et
al., 1989; Longhurst, 1991). ††††††† Les rťponses du systŤme aux perturbations transitoires sont
souvent complexes et non-linťaires. Des bilans
fiables sur le long terme ne peuvent Ítre calculťs si líon manque
de donnťes sur les variances et covariances ŗ petite ťchelle de temps,
en particulier aux ťchelles de quelques heures ŗ quelques jours. ††††††††††† En rťgime ocťanique,
les augmentations ťpisodiques de la biomasse phytoplanctonique reprťsentent
une fraction importante de la production primaire annuelle (Platt et
al., 1989) et donc de líexportation de matiŤre organique vers le fond
des ocťans. Les mťlanges turbulents induits par le vent sont un exemple
díťvťnements ťpisodiques qui peuvent modifier la dynamique et la composition
de líťcosystŤme pťlagique. Cette turbulence peut induire des transports
impulsionnels de sels nutritifs profonds vers la couche euphotique,
une augmentation de la biomasse et de la production phytoplanctonique,
des variations de composition des communautťs phyto- et zooplanctonique,
et une stimulation des flux verticaux de particules biogťniques (e.g. Haury et al., 1990; Marra et al., 1990; KiÝrboe, 1993). Les
flux verticaux de particules biogťniques sont aussi stimulťs par díautres
phťnomŤnes transitoires, tels que le dťveloppement rapide díessaims
de salpes (filtreurs gťlatineux) qui sont capables de consommer le
stock de petites cellules phytoplanctoniques et produisent de grandes
pelotes fťcales sťdimentant ŗ vitesse ťlevťe.
††††††††††† Díautre part, les pťriodes de transition saisonniŤre,
qui caractťrisent les zones tempťrťes, paraissent particuliŤrement
critiques car elles conditionnent
la variabilitť interannuelle. Ces pťriodes apparaissent gouvernťes
par la combinaison et la succession de processus rapides, de líordre
de quelques heures ŗ quelques jours. Leur ťtude doit Ítre menťe sur
plusieurs semaines consťcutives afin de bien apprťhender líeffet de
ses interactions et couplages, en particulier sur la succession des
espŤces dominantes qui peut Ítre brutale ou progressive. ††††††††††† ††††††††††† Par exemple, les observations ŗ haute frťquence rťalisťes
sur une pťriode díun mois dans la zone centrale de la mer Ligure
pendant la campagne DYNAPROC (mai 1995) ont permis de saisir
líimpact de coups de vent sur la dynamique de líťcosystŤme ŗ líťchelle
de quelques heures ŗ quelques jours, mais aussi leur rťpercussion
sur la transition saisonniŤre de la mťsotrophie ŗ líoligotrophie.
Cet impact a ťtť observť sur líťvolution de divers paramŤtres et
processus, tels que (1) la structure hydrologique et chimique dans
la couche euphotique avec, en particulier, un changement de líťlťment
nutritif limitant la croissance du phytoplancton et du rapport díutilisation
entre le carbone, líazote et líoxygŤne, (2) les biomasses, compositions
et distributions verticales du phytoplancton et du zooplancton, (3)
les activitťs bactťriennes, avec une ťvolution dans l'utilisation
de ressources carbonťes soit sous forme dissoute soit sous forme
particulaire, et (4) le stock de particules en suspension, leur temps
de rťsidence dans la couche euphotique, líamplitude et la composition
du flux vertical de matiŤre (Schmidt et al., 1997; Andersen & Prieur,
2000; Copin-Montťgut, 2000; Goutx et al., 2000; Pťrez et al., 2000;
Stemmann et al., 2000; Vidussi et al., 2000; Andersen et al., 2001a;
Van Wambeke et al., 2001). Les perturbations transitoires qui ont
eu lieu pendant la campagne ont díabord retardť la diminution de
la biomasse phytoplanctonique et donc líťvolution du systŤme vers
líoligotrophie, puis cette diminution a ťtť accťlťrťe sous l'effet
d'interactions complexes entre la disponibilitť en sels nutritifs,
la composition de la communautť phytoplanctonique et le broutage
du zooplancton. Le modŤle 1-D couplť physique-biologie appliquť aux
donnťes de la campagne a montrť que les contributions des processus
physiques et des processus biologiques ŗ la variation locale de la
biomasse du phytoplancton ťtaient du mÍme ordre de grandeur (Chifflet
et al., 2001). ††††††††††† Ces observations
montrent la nťcessitť díintťgrer ŗ l'ťchelle saisonniŤre, voire interannuelle,
les diffťrents processus physiques, chimiques et biologiques de rťgulation
se produisant ŗ de petites ťchelles spatio-temporelles et leur transfert ŗ líensemble
de la colonne d'eau. Ce type díťtude est indispensable pour ťtablir
un schťma cohťrent de la relation entre la production biologique de
surface et le flux díexportation en profondeur dont dťpend l'apport
de matiŤre organique au sťdiment.†
|
|
"Rťfťrences citťes
dans le
texte
Alldredge, A.L., Passow, U. & B.E. Logan, 1993. The abundance and significance of a class of large,††††† transparent organic particles in the ocean. Deep-Sea Res. II 40, 1131-1140 Amann, R.I., Ludwig, W. & K.H. Schleifer
, 1995. Phylogenetic
identification and in situ detection of individual microbial cells without
cultivation. Microbiol. Rev. 59,143-169. Amon R. M. W. & R. Benner, 1994. Rapid
cycling of high-molecular-weight dissolved organic matter in the ocean.
Nature 369, 549-552.† Amstrong, R. A., Lee, C., Hedges, J. I.
, Honjo, S.† & S. Wakeham.
Mineral ballast fluxes determine organic carbon fluxes to the deep ocean
and sediments. Nature
(en rťvision). Andersen, V. & L. Prieur, 2000. One-month study in the open NW Mediterranean
Sea (DYNAPROC experiment, May 1995): Overview of hydrobiogeochemical
structures and effects of wind events. Deep-Sea Res. I 47, 397-422. Andersen, V., Nival, P., Caparroy, P. & A.
Gubanova, 2001a. Zooplankton community during the transition from spring
bloom to oligotrophy in the open NW Mediterranean and effects of wind
events. 1 - Abundance and specific composition. J. Plankton Res. 23,
227-242. Andersen, V., Gubanova, A., Nival, P. & T.
Ruellet, 2001b. Zooplankton community during the transition from spring
bloom to oligotrophy in the open NW Mediterranean and effects of wind
events. 2 - Vertical distributionsand migrations. J. Plankton Res. 23,
243-261. Angel, M.V., 1989. Does mesopelagic biology
affect the vertical flux? In, Productivity of† the Ocean: Present and Past, Berger, W. H., Smetacek, V. S. & Wefer,
G. (eds.). John Wiley & Sons Limited, pp. 155-173. Azam F., 1998. Microbial control of oceanic
carbon flux : The plot thickens. Science† 280, 694-696.†††
Banse, K., 1994.† Grazing and zooplankton production as
key controls of phytoplankton production in the open ocean.† Oceanography 7, 13-20. Banse, K., 1995.† Zooplankton: Pivotal role in the control of
ocean production.† ICES J. mar. Sci. 52, 265-277. Becquevort S. & W.O. Smith Jr, 2001.† Aggregation, sedimentation and biodegradability of phytoplankton-derived
material during spring in the Ross Sea, Antarctica. Deep-Sea Res II 48,
4155-4178. Benitez-Nelson, C., Buesseler, K.O., Karl,
D.M. & J. Andrews, 2001. A time-series study of particulate matter
export in the North Pacific Subtropical Gyre based on 234Th: 238U
disequilibrium. Deep-Sea Res. I 48, 2595-2611. Buesseler, K.O., 1998. The decoupling
of production and particulate export in the surface ocean. Global Biogeochemical
Cycles 12, 297-310. Caillau, C., Belviso, S., Goutx, M. & A.
Bedo,, 1999. Particles flux in the Southern Ocean during Austral summer.
Mar. Ecol. Prog. Ser., 190 : 53-67 Carlson, D. J., Mayer, L. M., Brann, M.
L., & T. H. Mague, 1985. Binding of monomeric organic compounds to
macromolecular dissolved organic matter in sea water. Mar. Chem. 16:
141-153. Cauwet, G., 1994. HTCO method for dissolved
organic carbon analysis in seawater : influence of catalyst on blank
estimatiion. Mar. Chem. , 47 : 55-64. Chiarini, C., 1993. Micronecton et macroplancton en mer Ligure
(Mťditerranťe) : Distributions verticales et migrations nycthťmťrales
en automne. Universitť de GÍnes, Rapport de Stage de Recherches post-maÓtrise. Chifflet, M., Andersen, V., Prieur, L. & I.
Dekeyser, 2001. One-dimensional model of short-term dynamics of the pelagic
ecosystem in the NW Mediterranean Sea: effects of wind events. J. mar. Systems 30, 89-114. Christaki, U. & F. van Wambeke, 1995. Simulated bloom input in top-down manipulated
microcosms : comparative effect of zooflagellates, ciliates and copepods.
Aquat. Microbial Ecol. 9, 137 Ė 147. Chrůst, R.J. (ed.), 1991. Microbial Enzymes
in Aquatic Environments. Brock T.D. (Pub.), New York,† 317 pp.
Coale, K.H. & K.W. Bruland, 1985.† 234Th:238U
disequilibria within the California Current.† Limnol. Oceanogr. 30, 22-33. Copin-Montťgut, C., 2000. Consumption and production on scales of
a few days of inorganic carbon, nitrate and oxygen by the planktonic
community. Results of continuous measurements at the Dyfamed Station
in the northwestern Mediterranean Sea (May 1995). Deep-Sea Res. I 47,
447-477. Cottrell, M. & D. Kirchman, 2000a.
Natural assemblages of marine proteobacteria and members of the Cytophaga Ė Flavobacter
cluster consuming low- and high molecular weight dissolved organic matter,
Appl. Environ. Microbiol. 66 : 4, 1692-1697. Cottrell, M.T. & D.L. Kirchman, 2000b. Community composition of marine bacterioplankton
determined by 16S rRNA gene clone libraries and fluorescence in situ hybridisation. Appl. Env. Microbiol. 66, 5116-5122. DelbŤs, C, Moletta, R. & J.J. Godon,
2000. Monitoring of activity dynamics of an anaerobic digester bacterial
community using 16S rRNA polymerase chain reactionósingle-strand conformation
polymorphism analysis. Environ Microbiol. 2, 506-515. Dolan, J.R., 2000. Tintinnid ciliate diversity in the Mediterranean Sea:
longitudinal patterns related to water column structure in late spring-early
summer. Aquat. Microb. Ecol. 22,69-78. Fortier, L., Le FŤvre, J. & Legendre, L., 1994.† Export of biogenic carbon to fish and to the deep ocean:
the role of large planktonic microphages.† J. Plankton
Res. 16, 809-839. Fowler, S.W. & G.A. Knauer, 1986.† Role of large particles in the transport of elements
and organic compounds through the oceanic water column.† Prog. Oceanogr. 16, 147-194. Franks, P.J.S., 2001.† Phytoplankton blooms in fluctuating environment:
the roles of plankton response time scales and grazing.† J. Plankton
Res. 23, 1433-1441. Franqueville, C., 1971.† Macroplancton
profond (invertťbrťs) de la Mťditerranťe nord-occidentale.† Tethys
3, 11-56. FRONTAL, 1989. Rťsultats des missions "FRONTAL". Tome I:1984-1985. Tome II:1986-1988. Rapport interne, Villefranche-sur-mer, France.
Frost, B.W. & Franzen, N.C., 1992.† Grazing and iron limitation in the control of phytoplankton
stock and nutrient concentration: a chemostat analogue of the Pacific
equatorial upwelling zone.† Mar. Ecol. Prog. Ser. 83, 291-303. Gaspar, P., Grťgoris, Y. & J.-M. Lefevre, 1990.† A simple eddy kinetic energy model for simulations of
the oceanic vertical mixing: Tests at station Papa and long-term upper
ocean study site.† J. Geophys.
Res. 95, 16179-16193. Gonzalez, J.M., Simo, R., Massana,R.,
Covert, J.S., Casamayor, E.O., Pedros-Alio, C. & M. Ann Moran, 2000.
Bacterial community structure associated with a dimethylsulfoniopropionate-producing
North Atlantic algal bloom. Appl. Environ. Microbiol. 66, 4237-4246. Gorsky, G., Flood, P.R., Youngbluth, M.J.,
Picheral, M. & J.-M. Grisoni, 2000. Zooplankton Distribution in Four
Western Norwegian Fjords. Estuar. Coast. Shelf† Sci.
50, 135-141. Goutx, M., Momzikoff, A., Striby, L.,
Andersen, V., Marty, J.-C. & I. Vescovali, 2000. High frequency fluxes
of labile compounds in the central Ligurian Sea, northwestern Mediterranean.
Deep-Sea Res. I 47, 533-556. Goyet, C. & S.D. Hacker, 1992. Procedure
for calibration of a coulometric system used for total inorganic carbon
measurements of seawater. Mar. Chem.
38, 37-51. Grťgori, G., Citterio, S., Ghiani, A., Labra, M., Sgorbati, S. & M.
Denis, 2001. Resolution of viable and membrane-compromised bacteria in fresh water
and marine waters based on analytical flow cytometry and nucleic acid
double staining. Appl. Environ. Microbiol. 67, 4662-4670. Haury, L.R., McGowan, J.A. & Wiebe, P.H., 1978. Patterns and processes in time-space scales
of plankton distributions. In, Spatial pattern in plankton communities,
Steele, J. H. (ed.). Plenum Press, New York, pp. 277-327. Haury, L.R., Yamazaki, H. & E.C. Itsweire,
1990.† Effects of turbulent shear
flow on zooplankton distribution.† Deep-Sea
Res. 3, 447-461. Ittekott,V., Schűfer, P. Honjo, S., Depetris,
P.J.,(eds), 1996. Particule flux in the ocean, Scope 57. Wiley, New York. JŁrgens K. & H. GŁde, 1994. The potential importance of grazing-resistant
bacteria in planktonic systems. Mar. Ecol. Prog. Ser. 112, 169-188.
Karner M. & G.J. Herndl, 1992. Extracellular
enzymatic activity and secondary production in free-living and marine-snow-associated
bacteria. Mar Biol 113, 341-347. Kieber, R.J., Hydro, L.H. & P.J. Seaton,
1997. Photooxidation of triglycerides and fatty acids in seawater: Implication
toward the formation of marine humic substances. Limnol. Oceanogr. 42, 1454-1462. KiÝrboe, T., 1993.† Turbulence, phytoplankton cell size, and the structure
of pelagic food webs.† Adv. Mar. Biol. 29, 1-72. Klein, P. & B. Coste, 1984.† Effects of wind stress variability on nutrient transport
into the mixed layer.† Deep-Sea
Res. 31, 21-37. Klug, M.J. & J.M. Tiedje, 1993. Response of microbial communities to changing
environmental conditions : chemical and physiological approaches. In,
Trends in microbial ecology. R. Guerrero & C. Pedros-Alio (eds.),
pp. 371-386. Kuznetsova, M. & C. Lee, 2001. Enhanced extracellular enzymatic peptide
hydrolysis in the sea-surface microlayer. Mar. Chem.
73, 319-332. Lacroix, G., 1998. Etude de la variabilitť saisonniŤre et interannuelle
díun ťcosystŤme en Mer Ligure ŗ líaide díun modŤle couplť physique/biologie.
ThŤse de Doctorat en Sciences de líUniversitť de LiŤge et ThŤse de Doctorat
de líUniversitť Paris VI (Ocťanologie Biologique et Environnement Marin). Lacroix, G. & P. Nival, 1998. Influence of meteorological variability
on primary production dynamics in the Ligurian Sea (NW Med Sea) with
a 1D hydrodynamic/biological model. J. mar. Systems 16, 23-50. Lagadeuc, Y., Boulť, M. & J.J. Dodson,
1997. Effect of vertical mixing on the vertical distribution of copepods
in coastal waters. J. Plankton Res. 19, 1183-1204. Lampitt, R.S. & A.N. Antia, 1997.
Particle flux in deep seas: Regional characteristics and temporal variability.
Deep-Sea Res. I 44, 1377-1403. Langdon, C., 1984. Dissolved oxygen monitoring
system using a pulsed electrode design performance and evaluation. Deep-Sea
Res. Part A 31, 1357-1368. Lantoine, F. & J. Neveux, 1997. Spatial and seasonal variations in abundance
and spectral characteristics of phycoerythrins in the tropical Northeastern
Atlantic Ocean. Deep-Sea Res. I, 44, 223-246. Lebaron, P., Ghiglione, J.F. ,Fajon, C. & P.
Normand, 1998. Ecological implication of the diversity within a colony
morphotype as determined by PCR-RFLP. FEMS Microbiol.
Lett. 160, 137-143. Lťvy, M., Mťmery, L. & J.M. Andrť, 1998.† Simulation of primary production and export fluxes in
the Northwestern Mediterranean Sea.† J. mar. Res. 56, 197-238.
Long R.A. & F. Azam, 1996. Abundant
protein-containing particles in the sea. Aquat Microb Ecol 10, 213-221. Longhurst, A.R., 1991.† Role of the marine biosphere in the global
carbon cycle.† Limnol. Oceanogr.† 36, 1057-1526. Marine Zooplankton Colloquium 1, 1989.† Future
marine zooplankton research - a perspective.† Mar.
Ecol. Prog. Ser. 55, 197-206. Marine Zooplankton Colloquium 2, 2001.† Future
marine zooplankton research - a perspective.† Mar.
Ecol. Prog. Ser.† 222, 297-308. Marra, J., Bidigare, R.R. & T.D. Dickey,
1990.† Nutrients and mixing, chlorophyll
and phytoplankton growth.† Deep-Sea
Res. 37, 127-143. Martin, V., 1997. Etude par cytomťtrie en flux de la distribution
des populations phytoplanctoniques en Mťditerranťe. Mise en relation
avec la production mťtabolique de CO2 et comparaison avec
le Golfe du Saint Laurent. ThŤse de Doctorat de líUniversitť de la Mťditerranťe. Marty, J.C., Chiavťrini, J., Pizay, M.D. & B. Avril, 2002. Seasonal and interannual dynamics of nutrients
and phytoplankton pigments in the Western Mediterranean Sea at the DYFAMED
time-series station (1991-1999). Deep Sea Res. II 49 (12) (sous presse) Ohman, M.D., Drits, A.V., Clarke, M.E. & S. Plourde, 1998.† Differential dormancy of co-occurring
copepods.† Deep-Sea Res. II 45,
1709-1740. Palumbo, A.V., Ferguson, R.L. & P.A.
Rublee, 1984. Size of suspended bacterial cells and association of heterotrophic
activity with size fractions of particles in estuarine and coastal waters. Appl. Environ. Microbiol. 48, 157-164. Pťrez, M.T., Dolan, J.R., Vidussi, F. & E. Fukai, 2000. Diel vertical distribution of planktonic
ciliates within the surface layer of the NW Mediterranean (May 1995).
Deep-Sea Res. I 47, 479-503.
Platt, T., Harrison, W.G.,† Lewis, M.R., Li, W.K.W., Sathyendranath, S.,
Smith, R.E. & Vťzina, A.F., 1989.† Biological
production of the oceans : the case for a consensus.† Mar. Ecol.
Prog. Ser. 52 , 77-88. Raimbault, P., Pouvesle, W., Sempťrť, R., Diaz, F. & N. Garcia,
1999a. A simple
procedure for simultaneous analysis of total and dissolved organic forms
of carbon, nitrogen and phosphorus in seawater using the wet-oxidation
technic. Mar. Chem. 66, 161-169. Raimbault, P., Diaz, F. & B. Boudjellal,
1999b. Simultaneous determination of particulate forms of carbon, nitrogen
and phosphorus collected on filters using a semi-automatic wet-oxidation
procedure. Mar. Ecol. Progr. Ser. 180, 289-295. Raimbault P., Slawyk G., Boudjellal B.,
Coatanoan C., Conan P., Coste B., Garcia N., Moutin T. & M. Pujo-Pay,
1999c. Biomass, new production and export in the equatorial Pacific at
150įW: Evidence for intense nitrogen recycling. J. Geophys. Res. 104,
3341-3356. Raimbault P., Slawyk G. & N. Garcia,
2000. Comparison between chemical and isotopic measurements of biological
nitrate utilization: further evidence of low new production levels in
the equatorial Pacific. Mar. Biol., 136, 1147-115. Richardot M., Debroas, D., Thouvenot,
A., Romagoux, J.C., Berthon, J.L. & J. Devaux 1999. Proteolytic and
glycolytic activities in size-fractionated surface water samples from
an oilgotrophic reservoir in relation to plankton communities. Aquat.
Sci. 61, 279-292. Riemann, L., Steward, G.F. & F. Azam,
2000. Dynamics of bacterial community composition and activity during
a mesocosm diatom bloom. Appl. Environ. Microbiol. 66,
578-587. Sardou, J., Etienne, M. & Andersen, V., 1996.† Seasonal abundance and vertical distributions of macroplankton
and micronekton in the Northwestern Mediterranean Sea.† Oc. Acta 19, 645-656. Schmidt, S., Nival, P., Reyss, J.-L.,
Baker, M. & P. Buat-Mťnard, 1992. Relation between Th-234 scavenging
and zooplankton biomass in Mediterranean surface waters. Oceanol. Acta
15, 227-231. Schmidt, S., Reyss, J.-L. & V. Andersen,
1997. Residence times of particles in NW Mediterranean surface waters
estimated from 234Th / 238U disequilibria. Radioprotection - Colloques, 32, C2, 177-181. Slawyk G. & P. Raimbault,1995. A simple procedure for the simultaneous recovery
of dissolved inorganic and organic nitrogen in 15N-tracer experiments
on oceanic waters improving the mass balance. Mar. Ecol. Prog. Ser. 124,
289-299. Slawyk, G., Raimbault, P. & N. Garcia,
2000. Use of 15N to measure dissolved organic nitrogen release
by marine phytoplankton (reply to comment by Bronk and ward). Limnol.
Oceanogr. 45, 1884-1886. Smith, D.C., Steward, G.F. Long, R.A. & F.
Azam, 1995. Bacterial mediation of carbon fluxes during diatom bloom
in a mesocosm. Deep Sea Res. II 42, 75-97. Steele, J.H. & Henderson, E.W., 1992.† The
role of predation in plankton models.† J.
Plankton Res. 14, 157-172. Stemmann, L., Picheral, M. & G. Gorsky,
2000. Diel changes in the vertical distribution of suspended particulate
matter in the NW Mediterranean Sea investigated with the Underwater Video
Profiler. Deep-Sea Res. I 47, 505-531.
Tamburini C., Garcin J., Ragot M. & A.
Bianchi A., 2002. Biopolymer hydrolysis and bacterial production under
ambient hydrostatic pressure through a 2000 m water column in the NW
Mediterranean. Deep-sea Res. II, 49 (12) (sous presse). Touratier, F., Field, J. & C. Moloney,
2001. A stochiometric model relating growth substrate quality (C:N:P
ratios) to N:P ratios in the poducts of heterotrophic release and excretion. Ecol. Model. 139, 265 Ė 291. Trťguer, P. & P. Le Corre, 1975, Manuel d'analyse des sels
nutritifs dans l'eau de mer (Utilisation de l'AutoAnalyser II Technicon&).† Brest,† Laboratoire
d'Ocťanographie Chimique, Rapport interne, 110 pp. Van Wambeke, F., Goutx, M., Striby, L.,
Sempťrť, R. & F. Vidussi, 2001.† Bacterial
dynamics during the transition from spring bloom to oligotrophy in the
northwestern Mediterranean Sea: relationships with particulate detritus
and dissolved organic matter.† Mar. Ecol.
Prog. Ser 212, 89-105. Veldhuis M.J.W., Kraay G.W & K. Timmermans,
2001. Cell
death in phytoplankton: correlation between changes in membrane permeability,
photosynthetic activity, pigmentation and growth. J. Phycol. 36, 167-177. Verity, P.G. & Smetacek, V., 1996.† Organism
life cycles, predation, and the structure of marine pelagic ecosystems.† Mar. Ecol. Prog. Ser. 130, 277-293. Vidussi F., Claustre H., Bustillos-Guzman
J., Cailliau C. and J.C. Marty, 1996. Rapid HPLC method for determination
of phytoplankton chemotaxonomic pigments: separation of chlorophyll a
from divinyl chlorophyll a and zeaxanthin from lutein.† J.
Plankton Res. 18, 2377-2382. Vidussi, F., Marty, J.-C. & J. Chiavťrini,
2000. Phytoplankton pigment variations during the transition from spring
bloom to oligotrophy in the Mediterranean Sea. Deep-Sea Res. I 47, 423-445. Wakeham, S. G., & C. Lee, 1993. Production,
transport, and alteration of particulate organic matter in the marine
watercolumn. In : (M.H. Engel and S.A. Macko, Eds) Organic Geochemistry,
principles and applications, 145-169. Weeks, A., Conte, M. H., Harris, R. P.,
Bedo, A., Bellan, I., Burkill, P. H., Edwards, E. S., Harbour, D. S.,
Kennedy, H., Llewellyn, C., Mantoura, R.F.C., Morales, C. E., Pomroy,
A. J., & C. M. Turley, 1993. The physical and chemical environment
and changes in communauty structure associated with bloom evolution:
the Joint Global Flux Study North Atlantic Bloom Experiment. Deep-Sea
Research II, 40, 347-368. Williams P.J. LEB. & N.W. Jenkinson,
1982. A transportable microprocessor-controlled precise Winkler titration
suitable for field station and shipboard use. Limnol.
Oceanogr. 27, 576-585. |
|