L’opération MELISSA s’est déroulée en 2004. Compte tenu du budget finalement alloué en 2004 (15 k€ pour 11 campagnes au site DYFAMED, ce qui est très inférieur au soutien sollicité au départ), les objectifs scientifiques ont dû être recentrés sur les activités suivantes :
-
Etudes des processus de dissolution du matériel atmosphérique
-
Etudes de production nouvelle et régénérée, C, N, P et fixation d’azote moléculaire
-
Niveaux de fer dissous
Les travaux en mer étaient prévus pour 12 campagnes mensuelles, 11 se sont effectivement déroulées (manquent septembre et novembre, mais une sortie supplémentaire a été effectuées en février). ’arguments scientifiques, la motivation principale qui a conduit à sélectionner ces études parmi d’autres a été le souci de préserver les conditions de travail d’étudiants et de post-docs engagés sur MELISSA.
Planktonic
growth depends among others on the availability of several chemical species. As
the surface layer of the western Mediterranean is subjected to seasonal
succession of nutrient inputs, the main objective of the present project is to
better understand causal relationships between chemical constraints and the
functioning of the trophic chain. Climatic changes partly determine the
evolutions of such constraints, which have consequences on carbon fluxes. In
this way, this project will provide data for the study of retroaction of
chemical limitation of the biological production on the fate of CO2,
and, thus, on climatic evolutions.
The
present project will integrate time-series data acquired at the DYFAMED site
(central Ligurian Sea) since 1991. The seasonal evolution of N/P ratios clearly
indicate a succession of the limiting factor between N and P. This seasonal
pattern is linked to the temporal variability of nutrient inputs in the western
Mediterranean Sea. Certain phenomena are not well identified, and the
characterisation of seasonal features is not easy (e.g., the rapid and early P
to N limitation at the end of the stratification period).
It is
proposed here to integrate multidisciplinary approaches: nutrient inputs and
their availability in the euphotic zone, concomitant evolutions of the trophic
system (impact of a given limitation on planktonic dynamics), and its
consequences on carbon fluxes. These data will be simultaneously taken into
account, which will permit to better understand the involved processes, from
the input of nutrient to the exported fluxes. Reciprocally, measurements of
microbial respiration will provide data on the retroaction of seasonal
limitations on pCO2 variations.
Experimental
and field data (processes of stimulation of the biological production, of
trophic transfers, of recycling, etc.) will provide informations for the
conception and development of process models.
As far
as environmental evolutions, in particular anthropogenic perturbations, are
likely to influence the C:N:P:Si ratios and the abundance of dissolved iron,
the results acquired after three years in the present project (including in
vitro and in situ fertilising experiments), completed by the DYFAMED
time-series, will feed predictive models.
The
characterisation of seasonal situations implies the understanding of the impact
of events (e.g., a significant atmospheric event) and assimilation processes.
The different fields on which studies will focus are:
- Atmospheric
inputs
Atmospheric
fluxes of N, P, Si and Fe will be characterised and quantified, and their
temporal variability (event/season/interannual) will be assessed.
Solubilisation kinetics will be studied, since the physico-chemical form under
which nutrients are introduced into the surface layer determines their
assimilation by biota.
- Stocks
and fluxes of biogenic elements
The
seasonal succession of nutrient concentrations will be measured in surface
waters together with matter fluxes. Ultra-sensitive analytical methods will be
used for the stratification period. Vertical profiles will be measured in the
water column at the DYFAMED site to assess the distribution of dissolved
mineral and organic stoks of N, P, Si, Fe and C.
- Production
Primary
production (dissolved + particulate) monthly data from the Service
d’Observation DYFAMED will be used, as well as specific measurements during 3
campaigns in 2004. New and regenerated production will be measured. Bacterial
activity will be measured in the surface water column (0-200 m). The temporal
variability of exported carbon fluxes will be studied through monthly data from
the Service d’Observation DYFAMED, and through specific measurements during the
2004 campaigns.
- Biomass
Seasonal
variations of phytoplanktonic biomass (total + specific) will be monthly
measured, and also during the 2004 campaigns. Seasonal variations of
picoplankton abundance will be studied by flux cytometry, and the genetic
diversity of populations will be studied in the water column.
- DOM-TEP
The
biogeochemical significance of dissolved organic matter (DOM) will be addressed
by chemically characterising its high molecular weight (HWM) fraction. The role
of different factors that could contribute to seasonal and interannual
variations in the production and sinks of this material such as variations in
sources (algal and bacterial) in the availability of inorganic nutrients, and
the ability of different bacterial species to hydrolyse specific dissolved
biomolecules at different depths will be investigated. Transparent Exopolymeric
Particles (TEP), which may be a significant source of carbon, and may be a
favourable support for bacterial development, will be studied.
- Assimilation
processes
Although
the observed P-limitation in several oceanic zones is due to the biological
fixation of N2, the quantification of this process has not yet been
carried out in the Mediterranean. This process will be studied here by 15N
measurements. Genetics studies (characterisation of the genus nifH) will
be complementary. The study of the N2 fixation process will be
carried out in relation with the quantification of other nutrient fluxes, in
particular P and Fe.
The
relation between nutrient limitation and microbial population diversity will be
assessed. This point will be studied through experiments based upon the concept
of competitive assimilation of phosphate, in a situation of P-limitation (i.e.,
oligotrophic conditions).
The
efficiency of bacterial growth will be estimated in the surface layer (0-200 m)
by measuring the microbial production and respiration. Heterotrophic metabolism
will be studied in relation with COP and DOC fluxes.
- Fertilising
experiments
On the
basis of nutrient atmospheric fluxes measured at the Cap Ferrat sampling
station, fertilising experiments will be carried out at the DYFAMED site: LET
GO apparatuses will be spiked and incubated in situ with nutrient
concentrations calculated from wet atmospheric events. The role of mineral dust
will be examined too (Fe, Si, P). These works will be completed by in vitro
experiments.
- Modelling
The processes involved in the successive chemical limitations will
be modelled. The data acquired in this project will allow to follow the
evolutions of the soichiometric ratios in the exported material. On the basis
of stoichiometric variations, the vertical export of carbon out of the euphotic
zone will be constrained by values of i) primary production, ii) heterotrophic
respiration, iii) net carbon fixation, iv) chemical composition of the
dissolved and particulate exported material (C:N or C:P).
The impact of external inputs of nutrients on P:Si:N:C
ratios will be modelled in order to forecast the trends of a system driven by
climate change and anthropogenic activities (e.g., what are the consequences of
the evolution of molar ratios on the planktonic populations at the decenal
scale ?). Neural network models will be used in order to model stochastic
processes of the ecosys
AVANT-PROPOS
Ce projet a déjà été soumis au mois de février.
Les rapports ne remettent pas en cause la qualité scientifique
du projet, mais se rejoignent sur trois points majeurs :
1) Le plan de travail, la description des
protocoles, le coût des travaux, le temps-bateau, etc., doivent être
explicités ;
2) Les coûts globaux doivent être réduits ;
3) L’expérience de simulation d’un processus
naturel de fertilisation des eaux de surface doit être découplée
de la partie « caractérisation des configurations trophiques
saisonnières » du projet, c’est-à-dire tout ce qui concerne la
caractérisation des états trophiques saisonniers ;
Le projet
a en conséquence subi les modifications suivantes :
- Les
aspects pratiques du projets ont été détaillés. Chaque contributeur
a précisé ses interventions en termes de protocoles et de coûts.
- Le
coût total du projet a été sensiblement diminué : le montant
total s’élève à 183 411 €, pour une durée de 3 ans.
- L’expérience
de simulation en milieu naturel a été supprimée. Elle ne constitue
pas l’aboutissement des études de caractérisation des limitations
chimiques et de leurs conséquences sur le réseau trophique (qui
constituent en soi une question scientifique à part entière),
mais une vérification in situ des connaissances acquises
sur les successions temporelles des facteurs limitants et des états
trophiques associés. Une telle expérience fera éventuellement
l’objet d’un deuxième projet, complémentaire de celui-ci.
Par ailleurs, la présentation
de la problématique scientifique a été raccourcie.
PROBLEMATIQUE SCIENTIFIQUE
Limitations chimiques de
la production biologique
La connaissance des facteurs
qui limitent la production primaire, leurs successions temporelles et leurs
interdépendances constituent une question majeure de l’océanographie, comme
l’indiquent les actions entreprises dans le cadre des programmes IGBP, en
particulier JGOFS. Les sources d'approvisionnement des eaux marines de surface
en substances nutritives sont aussi bien internes (mouvements hydrologiques,
processus de recyclage) qu'externes (échanges aux détroits, cours d'eau,
atmosphère). La disponibilité des nutriments est régulée par un grand nombre de
paramètres et de processus physiques, chimiques et biologiques.
Les flux atmosphériques sont susceptibles d'engendrer, par
dissémination à l'échelle pélagique d'apports nutritifs externes (Jickells,
1995; Guerzoni et al., 1999; Migon et al., 2001; Guieu et al.,
2002a), un accroissement net de la biomasse. Les forçages environnementaux, et
notamment les apports atmosphériques, pourraient ainsi contraindre à
différentes échelles de temps les stocks de substances nutritives de la couche
euphotique, du moins à certaines périodes de l’année.
La Méditerranée occidentale est un site privilégié pour l'étude
des réponses marines aux perturbations chimiques de l'environnement (Béthoux et
al., 1990a, b, 1999; Martin & Milliman, 1997). Les apports
atmosphériques de nutriments pourraient au moins épisodiquement se révéler une
source privilégiée pour le milieu pélagique, quand les eaux de surface du large
sont stratifiées et isolées des eaux sous-jacentes (Migon et al., 1989;
Bergametti et al., 1992; Migon & Sandroni, 1999).
Quels sont les facteurs chimiques qui contrôlent la dynamique
trophique dans la couche de surface de la Méditerranée ? Dans les eaux méditerranéennes,
la concentration en nitrate est en moyenne 20 fois supérieure à la
concentration en phosphate. Le rapport moléculaire NO3-/PO43-
est en moyenne de 11 dans les cours d’eau (Meybeck, 1982), et de 22 dans les
eaux profondes de Méditerranée (Béthoux et al., 1998). Ce déséquilibre
pourrait s’expliquer par l’assimilation de N2 par certains
microorganismes marins (diazotrophie), la dissolution de l’azote moléculaire
atmosphérique constituant théoriquement un stock inépuisable de N. Des travaux
récents basés sur la mesure des rapports isotopiques naturels de l'azote
confirment cette hypothèse aussi bien en Méditerranée orientale (Sachs &
Repeta, 1999) qu'en Méditerranée occidentale (Kerhervé et al., 2001). En
Méditerranée, toutefois, les microorganismes capables de fixer N2
restent à identifier.
Au contraire de N, l’apport de P dépend essentiellement des
apports de cours d’eau (Froelich, 1988; Moutin et al., 1998), des
apports atmosphériques d’origine continentale, en particulier saharienne
(Bergametti et al., 1992; Guerzoni et al., 1999; Ridame &
Guieu, 2002), ou anthropique (Migon & Sandroni, 1999; Migon et al.,
2001). Ceci pourrait renforcer l’idée que P soit un facteur de limitation de la
production primaire en Méditerranée (Béthoux et al., 1998; Thingstad et
al., 1998).
Les eaux de surface méditerranéennes sont globalement
oligotrophes et limitées par P, mais certains paramètres environnementaux
présentent une variabilité saisonnière marquée. On peut admettre que le
phytoplancton et les bactéries hétérotrophes dans la couche euphotique soient
limitées par P pendant la période de stratification des eaux, alors que
d’autres espèces chimiques peuvent être des facteurs limitants en d’autres
périodes. Par exemple, le silicium (Si) peut contrôler la composition des
assemblages phytoplanctoniques quand sa disponibilité par rapport à N et P est
faible (Del Amo et al., 1997). Enfin, certains métaux sont nécessaires à
la croissance phytoplanctonique, en particulier le fer (Fe). En l’absence
d’apport atmosphérique significatif au moment où la colonne d’eau commence à se
stratifier, les concentrations en surface s’appauvrissent à cause de l’activité
biologique et peuvent devenir très faibles, voire limitantes. Les apports
atmosphériques sont une source majeure de fer, plus particulièrement en période
de stratification, où les eaux de surface sont isolées des eaux profondes
(Guieu et al., 2002). En Méditerranée, des apports atmosphériques pulsés
comme les événements sahariens sont très riches en Fe et pourraient épisodiquement
faire passer la colonne d’eau d’un état limité à un état excédentaire. S’il a
été démontré que les profils de concentrations sur les 40 premiers mètres de la
colonne d’eau sont effectivement contrôlés par les apports atmosphériques
sahariens (Guieu et al., 2002b), les données restent aujourd’hui rares
et parcellaires.
La solubilité et la forme physico-chimique des apports de Fe
sont également des paramètres déterminants pour l’évaluation du stock de Fe
biodisponible en fonction des cycles saisonniers. En Méditerranée, au site
DYFAMED, il a été montré que les concentrations de fer dissous dans la couche
de surface pouvaient attendre des valeurs extrêmement faibles (< 0,1 nM), au
moment où la couche de surface se stratifie et où la floraison printanière se
met en place (Sarthou & Jeandel,
2001). Le fer régénéré par broutage zooplanctonique
pourrait également être utilisé par le phytoplancton (Hutchins et al., 1993; Hutchins & Bruland, 1994).
Cependant, la question du facteur limitant la production
biologique n’est pas aussi simple, et le concept d’une succession de
limitations s’impose. La limitation pourrait par exemple passer de N à P et
vice-versa selon les conditions trophiques (Dolan et al., 1995; Klein et
al., 1997), comme le suggèrent les rapports N/P obtenus à partir des séries
temporelles du site DYFAMED (Marty et al., 2002). Par ailleurs, il a été
montré dans certaines zones océaniques (Atlantique tropical, Méditerranée) que
l'absorption de nitrate par le phytoplancton est activée par l'addition de
phosphate (Raimbault & Pujo-Pay, 1993; Diaz et al., 2001; Moutin
& Raimbault, 2002), et les apports de phosphate pourraient quant à eux
favoriser la fixation d’azote atmosphérique par les cyanobactéries dominantes
en période oligotrophe (Bonin et al., 1989).
Conséquences : cycle du carbone,
stoechiométrie, réseau trophique, diversité biologique
Dans des
conditions de faible (ou très faible) disponibilité (e.g. la période estivale
en Méditerranée), le phosphate est assimilé par les organismes de petite
taille, qui ont une forte affinité pour PO43-. Le réseau
trophique est dans ce cas essentiellement de type microbien, c’est-à-dire qu’il
est caractérisé par la compétition entre les algues de petite taille et les
bactéries hétérotrophes. C’est pourquoi on observe une nette dominance des
populations picoplanctoniques dans les milieux oligotrophes.
Dans des conditions de meilleure disponibilité (e.g., la
période printanière en Méditerranée), le phosphate est assimilé par des
organismes de plus grande taille, qui ont une moins forte affinité pour PO43-.
On a alors des chaînes trophiques plus étendues (puisqu’il y a davantage de
phosphate à partager), pouvant aller dans certaines situations (sporadiquement
en milieu côtier) jusqu’à l’eutrophisation.
Quelles sont les implications pour le cycle du carbone ? En
configuration d’oligotrophie, le phosphate est fixé de manière compétitive
entre les bactéries hétérotrophes et le picophytoplancton. Une augmentation de
la production bactérienne a été effectivement observée après un enrichissement
en phosphate des eaux de surface en période de stratification en Méditerranée
occidentale (Thingstad et al., 1998). Par conséquent, selon le modèle de
chaîne trophique représenté en Figure 1 (Thingstad et al., 1997),
l’excrétion de carbone organique dissous (COD) peut être supérieure à ce que
les bactéries peuvent consommer (dans l’hypothèse où leur croissance est
régulée par la disponibilité en P). Ce schéma conduit à une accumulation de COD
dans les eaux de surface et à son exportation, effectivement observés au site
DYFAMED (Avril, 2002). En cas de plus grande disponibilité du phosphate,
les chaînes trophiques plus étendues conduisent à une exportation du carbone
vers les couches profondes par sédimentation des particules (exportation de
carbone organique particulaire (COP).
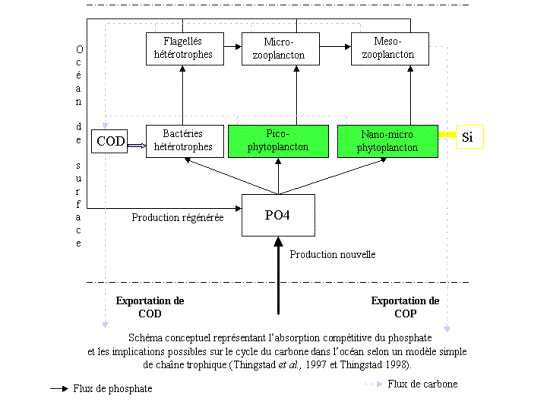
Figure 1: Model of
P-regulated trophic chain
Ce modèle suggère que le passage de
l’oligotrophie vers l’eutrophie se traduit par le passage d’un système caractérisé
par l’exportation de COD vers un système d’exportation de carbone via la
sédimentation des particules.
Quelle relation entre la limitation
par les nutriments et la diversité des populations microbiennes ?
Quel est l'effet des
gradients de sels nutritifs sur la diversité génétique des populations du
picoplancton ? Il a pu être montré que, sur la verticale (entre 0 et 150
m), il existe une stratification des populations, avec des génotypes de
profondeur génétiquement distincts des génotypes de surface. Mais cette
stratification semble plus liée au gradient vertical de lumière qu'à celui de
nutriments. En effet, les mêmes génotypes (de profondeur ou de surface) se
retrouvent partout en Méditerranée, des zones mésotrophes de l'entrée de la
Méditerranée aux eaux ultra-oligotrophes au large de la Crète, indiquant une
homogénéisation horizontale des populations de Prochlorococcus (Garczarek
et al., 2001).
Transfert de la matière
Le devenir de la matière
exogène dépend directement de la production biologique, donc de la
disponibilité des nutriments. En dehors des mélanges hydrologiques verticaux
(Copin-Montégut & Avril, 1993), localisés dans l’espace et dans le temps,
la matière dissoute est évacuée de la couche marine de surface à travers son
assimilation par le phytoplancton ou, beaucoup moins quantativement, par
adsorption sur des particules ou des bactéries (Fowle & Fein, 2000). La
vitesse de chute des particules lithogéniques d’origine atmosphérique, étant
négligeable dans l’eau de mer (Bishop et al., 1977; Buat-Ménard et al.,
1989 ; Migon et al., 2002), leur transfert vertical se fait par
incorporation biologique dans les grosses particules organiques (Deuser et
al., 1983; Fowler et al., 1987; Fisher et al., 1991) et par
agrégation (Wells & Goldberg, 1994).
Ainsi, la structure saisonnière des flux de particules
traduit-elle à la fois la productivité du système (Marty et al., 1994;
Miquel et al., 1994) et l'efficacité du transfert vertical de la matière
d’origine atmosphérique (Buat-Ménard et al., 1989; Jickells et al.,
1998; Migon et al., 2002).
Le site DYFAMED voit se succéder au cours de l’année tous les
états trophiques qu’on peut rencontrer en Méditerranée hauturière : les
conditions hivernales sont caractérisées par un mélange vertical intense qui
remet à disposition les nutriments accumulés en profondeur, permettant ainsi
des floraisons phytoplanctoniques printanières; l’été est caractérisé par la
stratification des eaux du large, ce qui pourrait favoriser la voie
d’approvisionnement atmosphérique (Migon et al., 1989; Bergametti et
al., 1992; Migon & Sandroni, 1999); enfin, les coups de vents
interviennent épisodiquement (Andersen & Prieur, 2000; Vidussi et al.,
2000). Il faut aussi mentionner la stabilité interannuelle de la distribution
de la production planctonique observée au site DYFAMED (Marty et al.,
2000).
Les bactéries, par leurs capacités d'hydrolyse, utilisent les
formes particulaire et dissoute de la matière organique, différemment selon la
profondeur et selon la saison (selon l'état trophique), en relation avec les
caractéristiques des réseaux trophiques, jouant ainsi un rôle déterminant dans
le contrôle du flux particulaire (Goutx et al., 2000; van Wambeke et
al., 2001).
Certaines particules organiques non vivantes (hors pelotes
fécales, cadavres et neige marine) pourraient aussi jouer un rôle important
dans le transfert du carbone (Mari et al., 2001). La fraction colloïdale
de la matière organique dissoute (MOD) libérée par le phytoplancton et les
bactéries (exsudation active ou passive du phytoplancton) conduit par
coagulation spontanée à la formation de particules exopolymériques
transparentes (Transparent Exopolymeric Particles : TEP), très riches en
carbone. La taille de ces particules varie entre 1 et 100 µm. La période
d’abondance maximale des TEP est l’été (> 30 000 particules ml-1).
La formation de TEP favorise-t-elle la consommation de MOD par les bactéries ?
Des travaux préliminaires effectués au site DYFAMED ont montré qu’en été, la
matière organique des TEP a un rapport C:N de 39 et représente environ 30% du
COD (20 à 25 µmol C dans les TEP pour 80 µmol C pour la MOD ; Copin-Montégut
& Avril, 1993). Reste à savoir si le carbone des TEP est disponible pour
les organismes pélagiques.
Une situation de limitation conduit à une baisse de l’activité
microbienne, donc une moindre dégradation de la MOD. Cela devrait avoir pour
conséquence une augmentation de la concentration des TEP. Peut-on par
conséquent utiliser les TEP comme indicateurs de régime trophique ?
Les TEP pourraient avoir un impact sur les flux de carbone en
zone pélagique à deux niveaux : i) en stimulant le transport vertical de
matière par des processus de coagulation et de sédimentation (Jackson, 1995;
Logan et al., 1995); et ii) en étant une source de carbone pour un
certain nombre d’organismes, dont les bactéries (Decho & Moriarty, 1990;
Flood et al., 1992, Tranvik et al., 1993). Les TEP pourraient
représenter un mode de transport significatif pour le COD, c’est-à-dire une
voie alternative à la boucle microbienne (Mari, 1999).
Enfin, les TEP pouvant se former par coagulation de
polysaccharides qui dominent la fraction à haut poids moléculaire de la matière
organique dissoute (DOM), une fraction importante des stocks de DOM pourraient
être exportée via la formation des TEP (Gogou & Repeta, 2000).
La connaissance des mécanismes de
production et des transformations de la DOM qui conduisent à l'assimilation
microbienne ou à l'exportation du carbone en relation avec un type de
limitation chimique s'inscrit dans la problématique du présent projet. La
signification biogéochimique de la DOM peut être approchée par la
caractérisation chimique de sa fraction à haut poids moléculaire (HMW), qui
représente 25 à 35% de la DOM totale.
Les organismes zooplanctoniques, résultant eux-mêmes des
diverses limitations chimiques exercées sur le système, jouent un rôle
essentiel dans le transfert de la matière au sein des écosystèmes pélagiques,
déterminant par leur activité alimentaire une grande partie du devenir de la
production primaire et de la production microbienne au sens large (i.e.
incluant bactéries et protistes hétérotrophes). Le zooplancton participe en
effet i) à la reminéralisation de la matière organique (respiration,
excrétion), ii) au recyclage et au maintien du carbone dans la zone euphotique
(transfert vers les niveaux trophiques supérieurs), et iii) aux transferts vers
les couches marines profondes : migrations verticales, qui peuvent transporter
de grandes quantités de carbone organique ultérieurement décomposé et respiré
en grande profondeur (séquestration; Dam et al., 1995; Steinberg et
al., 2000), et production de pelotes fécales (Fowler & Knauer, 1986;
Alldredge & Silver, 1988). En outre, les organismes zooplanctoniques
constituent eux-mêmes un stock relativement important de carbone et jouent un
rôle tampon dans les transferts de matière. Il faut par conséquent s'interroger
sur l'effet cascade d'une limitation en éléments nutritifs sur le compartiment
zooplanctonique.
Enfin, la modélisation a montré à travers l'évolution des
rapports C:N:P que les processus d'excrétion hétérotrophes (bactériens et
zooplanctoniques) induisent des changements dans la limitation elle-même
(Touratier et al., 2001). Ceci met en évidence la réciprocité des
relations "limitation chimique-production" et, en l'occurrence, la
nécessité de prendre en compte la variabilité saisonnière des systèmes
d'excrétion pour mieux définir les successions saisonnières de limitation
chimique.
Tendances et évolution interannuelle du système
Plusieurs paramètres
contribuent, à des échelles de temps différentes, à l'évolution des systèmes
marins, et du système méditerranéen en particulier : les changements
climatiques, météorologiques (influence variable des apports atmosphériques de
poussières désertiques, évolution du régime pluvial), l'augmentation de
l'irradiation UV des eaux de surface, pression anthropique, etc. Compte tenu de
l'accroissement des activités humaines autour de la Méditerranée, les apports
(notamment atmosphériques) de certains éléments nutritifs au milieu hauturier
augmentent régulièrement. C’est le cas de P, dont les concentrations dans les
eaux profondes de Méditerranée occidentale ont augmenté de 0,5% par an au cours
des 40 dernières années (Béthoux et al., 1998). Cette augmentation
pourrait être reliée à une croissance des apports en surface de l’ordre de 3%
par an, ce qui conduirait, typiquement, à un doublement des niveaux de
concentration en moins de 25 ans. Bien que cela soit plus difficile à mesurer,
ceci est probablement aussi valable pour N (via les émissions
anthropiques de nitrates).
Par contre, les apports de Si ne dépendent pratiquement pas du
facteur anthropique. Dans un écosystème perturbé par les activités humaines,
les rapports Si:N et/ou Si:P diminuent (Figure 2). De telles conditions peuvent
entraîner une modification des facteurs de contrôle des diatomées (passage à
une limitation par Si), voire modifier profondément la nature des communautés
phytoplanctoniques (passage d’un système dominé par les diatomées vers un
système dominé par des flagellés non siliceux dont certains peuvent s’avérer
toxiques) (Smayda, 1990; Billen et al., 1991; Schöllhorn & Graneli,
1993). Les rapports N:Si:P observés dans le bassin occidental (1:21:22; Béthoux
et al., 1998) pourraient être la résultante (via assimilation
biologique) d’un déséquilibre des apports de nutriments à la couche productive.
Ceci pourrait refléter une situation épisodique de limitation par Si.
L’exportation de matière carbonée dépend fortement de la
dominance du premier niveau trophique par les diatomées ou, plus globalement,
les espèces de grande taille (Buesseler et al., 1998). Pour ces
organismes phytoplanctoniques, Si est effectivement le facteur limitant (Nelson
et al., 1995; Tréguer et al., 1995). Par suite, la perturbation
des abondances relatives de N, P et Si pourrait induire un changement des
espèces planctoniques dominantes dans l’océan. On assiste peut-être déjà, en
Méditerranée occidentale, à une évolution d’un système à dominance de diatomées
vers un système à majorité d’organismes non-siliceux (Béthoux et al.,
2002). Les séries temporelles acquises au site DYFAMED (1991-1999) semblent
d'ailleurs indiquer une augmentation de la biomasse phytoplanctonique totale
(Chl a), alors que les stocks de microplancton restent constants, voire
diminuent légèrement (Marty et al., 2002). D’autre part, la pêche en
Méditerranée stagne aux alentours de 1,1 106 tonnes par an depuis
une dizaine d’années (UNEP, 1996).
Ce point fait apparaître que la limitation de la production
biologique doit être considérée à diverses échelles de temps, et les situations
saisonnières sont elles-mêmes susceptibles d'évoluer à l'échelle décennale.
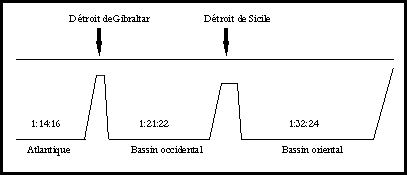
Figure 2: Present values of P/Si/N molar
ratios in deep Mediterranean and Atlantic waters (after Béthoux et al.,
2002).
OBJECTIFS
GENERAUX
Le développement planctonique
dépend de la conjonction favorable de conditions physiques, chimiques et
biologiques. Compte tenu de la variabilité saisonnière évidente des apports de
nutriments à la couche de surface en Méditerranée, l'objectif principal de ce
projet est de mieux comprendre les relations de causalité entre les forçages
chimiques apportés aux systèmes océaniques (c'est-à-dire, dans le cas présent,
la disponibilité des substances nutritives) et le fonctionnement du réseau
trophique. Ces forçages, dont les évolutions sont en partie déterminées par
les changements climatiques, agissent entre autres sur les flux de carbone. Par
conséquent, ce projet fournira en retour des éléments pour l'étude de la rétroaction
de la limitation chimique de la production sur le devenir du CO2, et
donc sur les évolutions climatiques.
Le présent projet propose d'intégrer les acquis des
observations au site DYFAMED depuis 1991. L'évolution saisonnière des
rapports N/P (Figure 3) indique clairement une succession du facteur limitant
entre N et P. Cette succession saisonnière est liée à la variabilité temporelle
caractéristique des apports de substances nutritives dans le bassin
méditerranéen occidental. On observe que certains phénomènes ne sont pas encore
clairement explicités et que la caractérisation des situations à l'échelle
saisonnière n'est pas aussi évidente qu'on pourrait le penser (e.g., le passage
rapide et précoce d'une limitation par P vers une limitation par N en fin de
période stratifiée).
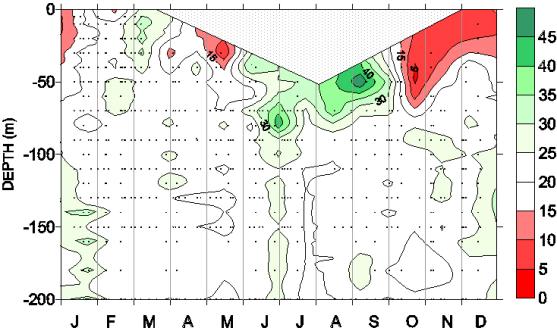
Figure 3: Seasonal variations of N/P
ratio in the 0-200 m layer for a composite year (1991-1999), after Marty et
al. (2002)
L'approche proposée intègre des investigations sur les apports
de substances nutritives et leur disponibilité dans la couche éclairée, les
évolutions concomitantes du système trophique (impact d'une limitation sur la
dynamique des populations planctoniques, des populations microbiennes au
zooplancton), et les conséquences sur les flux de carbone. La prise en compte
simultanée de ces données et de leur évolution temporelle permettra de mieux
appréhender les processus mis en jeu depuis la mise à disposition des éléments
chimiques jusqu’aux flux exportés. Réciproquement, les mesures de respiration
microbienne fourniront des éléments sur la rétroaction des limitations
saisonnières sur le climat, à travers les variations de pCO2. A ce
titre, le présent projet sera en étroite relation avec le projet ACTION.
Les données expérimentales et de terrain (processus de
stimulation de la production biologique, de transferts trophiques, de
recyclage, etc.) fourniront des éléments pour la conception et le développement
de modèles déterministes.
Enfin, les résultats acquis sur une durée de trois ans,
complétés par les séries temporelles du Service d’Observation (SO) DYFAMED
(évolutions environnementales, notamment le forçage anthropique, pouvant
influencer les rapports d'abondance entre les nutriments N, P, Si et le fer),
ainsi que les résultats des expériences de fertilisation (dopages d'enceintes
transparentes en milieu naturel), permettront d’alimenter des modèles
prédictifs.
Complémentarité des
approches
- Le présent projet est
essentiellement basé sur des observations de terrain, complétées par des travaux
in vitro.
- L'échelle de temps
est multiple : il s'agit de caractériser des configurations saisonnières,
ce qui implique la compréhension des processus, et donc d'intégrer l'échelle impulsionnelle.
A terme, cette approche ambitionne d'intégrer les données acquises au site
DYFAMED depuis 1991 et de fournir des éléments nouveaux pour la compréhension
des évolutions à l'échelle décennale.
- Pour la dimension
spatiale, le projet se positionne, du moins en ce qui concerne les
processus, à l'interface air-mer et la couche productive. Les acquis de
l'opération DYFAMED permettent la connexion avec les couches plus profondes.
Alldredge, A.L. & Silver, M.W. (1988) Characteristics, dynamics and
significance of marine snow. Progr. Oceanogr., 20, 41-82.
Andersen, T., 1997. Pelagic
nutrient cycles. Herbivores as sources and sinks. In: R. M. M.
Caldwell, G. Heldmaier, O. L. Lange, H. A. Mooney, E.-D. Schulze, U. Sommer
(Editors), Ecological Studies, Analysis and Synthesis, Springer, 129, 280 pp.
Andersen, V. & Nival, P.
(1988) Modèles d’écosystème pélagique des eaux côtières de la mer Ligure. In
: Océanographie Pélagique Méditerranéenne, H.J. Minas & P. Nival (eds), Oceanol.
Acta, SN 9, 211-217.
Andersen, V. & Prieur, L. (2000) One-month study in the open NW
Mediterranean Sea (DYNAPROC experiment, May 1995): overview of the
hydrobiogeochemical structures and effects of wind events. Deep-Sea Res. I, 47, 397-422.
Athias, V. (2001) Une méthode
d’estimation paramétrique non-linéaire pour modéliser les flux de matière dans
la colonne d’eau océanique, PhD Thesis, Univ. Paul Sabatier, Toulouse.
Avril, B. (2002) DOC dynamics in the northwestern
Mediterranean Sea. Deep-Sea Res. II, sous
presse.
Bergametti, G., Remoudaki, E., Losno, R., Steiner, E., Chatenet, B.
& Buat-Ménard, P. (1992) Source, transport and deposition of atmospheric
phosphorus over the northwestern Mediterranean. J. Atmos. Chem., 14,
501-513.
Béthoux, J.P., Courau, P., Nicolas, E. & Ruiz-Pino, D. (1990a) Trace
metal pollution in the Mediterranean Sea. Oceanol. Acta, 13, 4, 481-488.
Béthoux, J.P., Gentili, B., Morin, P., Nicolas, E., Pierre &
Ruiz-Pino, D. (1999) The Mediterranean Sea: a miniature ocean for climatic and
environmental studies and a key for the climatic functioning of the North
Atlantic. Progr. Oceanogr., 44, 131-146.
Béthoux, J.P., Gentili, B., Tailliez, D. & Raunet, J. (1990b)
Warming trend in the western Mediterranean deep water. Nature, 347,
660-662.
Béthoux, J.P., Morin, P., Chaumery, C., Connan, O., Gentili, B. &
Ruiz-Pino, D. (1998) Nutrients in the Mediterranean Sea, mass balance and
statistical analysis of concentrations with respect to environmental change. Mar.
Chem., 63, 155-169.
Béthoux, J.P., Morin, P. & Ruiz-Pino, D. (2002) Mediterranean
ecosystem: chemical evidence of change driven by human activity. Deep-Sea
Res. II, sous presse.
Billen G.L.C. & Meybeck M., (1991) N, P and Si retention along the
aquatic continuum from land to ocean. In: Ocean Margin Processes in Global
Change, R.F.C. Mantoura & R. Wollast (eds.), John Wiley & Sons Ltd, pp.
19-44.
Bishop, J.K.B., Edmond, J.M., Ketten, D.R., Bacon, M.P. & Silker,
W.B. (1977) The chemistry, biology and vertical flux of particulate matter from
the upper 400 m of the equatorial Atlantic Ocean. Deep-Sea Res., 24,
511-548.
Blain S., Guieu C., Claustre H., Leblanc
K., Moutin T., Newman S., Quéquiner B., Sarthou G. & Tréguer P. (2001)
Evidence of iron limitation in the North East Atlantic (Azore-Portugal), IGPB
Open Science Conference, Amterdam 10-13 July 2001.
Bonin, D.J., Bonin, M.C. & Berman, T. (1989)
Mise en évidence expérimentale des facteurs nutritifs limitants de la
production du micro-nanoplancton et de l’ultraplancton dans une eau côtière de
la Méditerranée orientale (Haïfa, Israël). Aquat. Sci., 51, 129-152.
Bowie, A.R., Maldonado, M.T.,
Frew, R.D., Croot, P.L., Achterberg, E.P., Mantoura, R.F.C., Worsfold, P.J.,
Law, C.S. & Boyd, P.W. (2001) The fate of added iron during a mesoscale
fertilisation experiment in the Southern Ocean. Deep-Sea Res. II, 48(11-12),
2703-2743.
Boyd et al. (2000) A mesoscale phytoplankton bloom in the polar
Southern Ocean stimulated by iron fertilization, Nature, 407, 695-702.
Buat-Ménard, P., Davies, J., Remoudaki, E., Miquel, J.C., Bergametti,
G., Lambert, C.E., Ezat, U., Quétel, C., La Rosa, J. & Fowler, S.W. (1989)
Non-steady state biological removal of atmospheric particles from Mediterranean
surface waters. Nature, 340, 131-134.
Buesseler, K.W. (1998) The decoupling of production and particulate
export in the surface ocean. Global Biogeochem. Cycles, 12, 297-310.
Chambard, V. (1999)
Vérification expérimentale du rôle fertilisateur du phosphore atmosphérique sur
le milieu marin. DEA Océanologie, Météorologie et Environnement, Univ. Paris 6,
30 pp.
Coale et al. (1996) A
massive phytoplankton bloom induced by an ecosystem scale iron fertilization
experiment in the equatorial Pacific Ocean, Nature, 383, 495-501.
Copin-Montégut, G. & Avril, B. (1993) Vertical distribution and
temporal variation of dissolved organic carbon in the north-western
Mediterranean Sea. Deep-Sea Res. I, 40, 10, 1963-1972.
Del Amo Y., Le Pape O., Tréguer P., Quéguiner B., Ménesguen A. &
Aminot A. (1997) Impacts of high-nitrate freshwater inputs on macrotidal
ecosystems. I. Seasonal evolution of nutrient limitation for the
diatom-dominated phytoplankton of the Bay of Brest (France). Mar. Ecol.
Prog. Ser., 161, 213-224.
Dam, H.G., Zhang, X., Butler, M. & Roman, M.R. (1995)
Mesozooplankton grazing and metabolism at the Equator in the Central Pacific:
implications for C and N fluxes. Deep-Sea Res. II, 42, 732-756.
Decho, A.W. & Moriarty, DJ.W. (1990) Bacterial exopolyme utilization
by a harpacticoid copepod: a methodology and results. Limnol. Oceanogr.,
35, 1039-1049.
Deuser, W.G., Brewer, P.G., Jickells, T.D. & Commeau, R.F. (1983)
Biological control of the removal of abiogenic particles from the surface
ocean. Science, 21, 388-391.
Diaz F. (2000) Evolution
saisonnière de la production primaire et des processus d’assimilation-
régénération de l’azote dans le golfe du Lion. Estimation d’un bilan de
carbone. Approches in situ et modélisation. PhD thesis, Univ. Aix-Marseille II, 341 pp.
Diaz, F., Raimbault, P., Garcia, N. & Moutin, T. (2001) Early
phosphorus limitation during spring in the Gulf of Lions. Mar. Ecol. Progr.
Ser., 211, 51-62.
Dolan, J.R., Thingstad, T.F. & Rassoulzadegan, F. (1995) Phosphate
transfer between size-fractions in Villefranche Bay (NW Mediterranean Sea),
France, in autumn 1992. Ophelia, 41, 71-85.
Echevin, V., Mortier, L. & Crépon, M. Analysis of the mesoscale
circulation in the North Western Mediterranean Sea, Annales Geophysicae,
soumis.
Elser, J.J. & Hassett, R.P. (1994) A stoichiometric analysis of the
zooplankton-phytoplankton interaction in marine and freshwater ecosystems. Nature,
370: 211-213.
Elser, J.J., Dobberfuhl, D.R., MacKay, N.A. & Schampel., J.H. (1996)
Organism size, life history, and N:P stoichiometry. Bioscience, 46:
674-684.
Eppley & Peterson (1979) Particulate organic matter flux and
planktonic new production in the deep ocean. Nature, 282, 677-680.
Faugeras, B., Lévy, M., Mémery, L., Blum, J., & Verron, J.
Estimating the parameters of a 1D biogeochemical model to assimilate date from
the DYFAMED station in the North Western Mediterranean Sea, J. Mar. Syst.,
soumis.
Fisher, N.S., Nolan, C.V. & Fowler, S.W. (1991) Scavenging and
retention of metals by zooplankton fecal pellets and marine snow. Deep-Sea
Res. I, 38, 10, 1261-1275.
Flood, P.R., Deibel, D. & Morris, C.C. (1992) Filtration of
colloidal melanin from sea water by plankton tunicates. Nature, 355,
630-632.
Fowle, D.A. & Fein, J.B. (2000) Experimental measurements of the
reversibility of metals-bacteria adsorption reactions. Chem. Geol., 168,
27-36.
Fowler, S.W., Buat-Ménard, P., Yokoyama, Y., Ballestra, S., Holm, E. and
Nguyen, H.V. (1987) Rapid removal of Chernobyl fallout from Mediterranean
surface waters by biological activity. Nature, 329, 56-58.
Fowler, S.W. & Knauer, G.A. (1986) Role of large particles in the
transport of elements and organic compounds through the wtare column. Progr.
Oceanogr., 16, 147-194.
Froelich, P.N. (1988) Kinetic control of dissolved phosphate in natural
rivers and estuaries : a primer on the phosphate buffer mechanism. Limnol.
Oceanogr., 33, 649-668.
Garczarek, L., Dufresne, A., Partensky, F. (2001), Use of a highly
variable gene (pcb) for studying the diversity of cultured and natural Prochlorococcus
genotypes. ASLO Aquatic Sciences 2001, Albuquerque, New Mexico, January 2001.
Gogou,
A. & Repeta, D.J. (2000). Chemical
characterization of dissolved and particulate organic matter in seawater: Do
particles pump DOM from the surface to the deep ocean ? Ocean Sciences Meeting,
San Antonio, Texas, January 2000.
Goutx, M., Momzikoff, A., Striby, L., Marty, J.C. & Vescovali, I.
(2000) High frequency fluxes of labile compounds in the Central Ligurian Sea;
Northwestern Mediterranean. Deep-Sea Res. I, 7, 533-556.
Guerzoni, S., Chester, R., Dulac, F., Herut, B., Loÿe-Pilot, M.D.,
Measures, C., Migon, C., Molinaroli, E., Moulin, C., Rossini, P., Saydam, C.,
Soudine, A. & Ziveri, P. (1999) The role of atmospheric deposition in the
biogeochemistry of the Mediterranean Sea. Progr. Oceanogr., 44, 147-190.
Guieu, C., Chester, R., Nimmo, M., Martin, J.M., Guerzoni, S., Nicolas,
E., Mateu, J. & Keyse, S. (1997) Atmospheric input of dissolved and
particulate metals to the northwestern Mediterranean,. Deep-Sea Res. II,
44, 3-4, 655-674.
Guieu C., Bozec Y., Blain S., Ridame C. , Sarthou G. &
Leblond N. (2002a) Impact of high Saharan dust inputs on dissolved iron
distributions in the Mediterranean Sea. Geophys. Res. Lett., sous presse.
Guieu, C., Loÿe-Pilot, M.D., Ridame, C. & Thomas, C. (2002b)
Chemical characterisation of the Saharan end-member; some biogeochemical
implications in the western Mediterranean. J. Geophys. Res., sous presse.
Hutchins, D.A., DiTullio, G.R. & Bruland, K.W. (1993) Iron and
regenerated production: Evidence for biological iron recycling in two marine
environments. Limnol. Oceanogr., 38, 6, 1242-1255.
Jackson, G.A. (1995) TEP and coagulation during a
mesocosm experiment. Deep-Sea Res. II, 42, 215-222.
Jickells, T.D. (1995) Atmospheric inputs of metals and
nutrients to the oceans: their magnitude and effects. Mar. Chem., 48,
199-214.
Jickells, T.D., Dorling, S., Deuser, W.G., Church,
T.M., Arimoto, R. & Prospero, J.M. (1998) Air-borne dust fluxes to a deep
water sediment trap in the Sargasso Sea. Global Biogeochem. Cycles, 12,
311-320.
Kamatani A. (1982) Dissolution rates of silica from diatoms decomposing
at various temperatures. Mar. Biol., 68, 91-96.
Karl, D.M., Dore, J.E., Lukas, R., Michaels, A.F., Bates, N.R. &
Knap, A. (2001) Building the long-term picture: the U.S. JGOFS time-series
programs. Oceanography, 14, 6-17.
Karl, D.M., Hebel, D.V., Björkman, K. & Letelier, R.M. (1998) The
role of dissolved organic matter release in the productivity of the
oligotrophic North Pacific Ocean. Limnol. Oceanogr., 43, 6, 1270-1286.
Karl, D.M. & Tien, G. (1992) MAGIC: a sensitive and precise method
for measuring dissolved phosphorus in aquatic environment. Limnol. Oceanogr.,
37, 1, 105-116.
Kerhervé, P., Minagawa, M., Heussner, S., Monaco,
A. (2001) Stable isotopes (13C/12C and 15N/14N)
in settling organic matter of the northwestern Mediteraen Sea: biogeochemical
implications. Oceanol. Acta, 24, 77-85.
Klein, C., Dolan, J.R. & Rassoulzadegan, F. (1997) Experimental
examination of the effects of rainwater on microbial communities in the surface
layer of the NW Mediterranean Sea. Mar. Ecol. Progr. Ser., 158, 41-50.
Kriest I. & Evans, J.T. (1999) Representing phytoplankton aggregates
in biogeochemical models, Deep-Sea Res.I, 46, 1841-1859.
Lévy, M., Mémery, L. & André, J.M. (1998a) Simulation of primary
production and export fluxes in the Northwestern Mediterranean Sea. J. Mar.
Res., 56, 197-238.
Lévy, M., Mémery, L. & Madec, G. (1998b) The onset of a bloom after
deep winter convection in the North Western Mediterranean Sea: mesoscale
process study with a primitive equation model, J. Mar. Syst., 16, 7-21.
Lévy, M., Mémery, L. & Madec, G. (1999) The onset of the spring
bloom in the MEDOC area: mesoscale spatial variability, Deep-Sea Res.,
1137-1160.
Lévy, M., Mémery, L. & Madec, G. (2000) Combined effects of
mesoscale processes and atmospheric high-frequency variability on the spring
bloom in the MEDIC area, Deep-Sea Res., 47, 27-53.
Logan, B.E., Passow, U., Alldredge, A.L., Grossart, H.P. & Simon, M.
(1995) Rapid formation and sedimentation of large aggregates is predictable
from coagulation rates (half-life) of transparent exopolymeric particles (TEP).
Deep-Sea Res.I, 42,
203-214.
Magro, V., Chambard, V., Chiaverini, J., Migon, C., Marty J.C., Stock,
A. & Perez, M. (2000) Experimental evidence of of marine surface water
fertilisation by atmospherically-transported phosphorus. 25th EGS
General Assembly, Nice (France), 25-29 avril 2000.
Mari, X. (1999) Carbon content and C:N ratio of transparent exopolymeric
particles (TEP) produced by bubbling exudates of diatoms. Mar. Ecol. Progr.
Ser., 183, 59-71.
Mari, X., Beauvais, S., Lemée, R. & Pedrotti, M.L. (2001)
Non-Redfield C:N ratio of transparent exopolymeric particles in the
northwestern Mediterranean Sea. Limnol. Oceanogr., 46, 7, 1831-1836.
Mac Neil, J.D., Jannasch, H.W., Dickey, T.,
McGillicuddy, D., Brzezinski, M., & Sakamoto, M. (1999) New chemical,
bio-optical and physical observations of upper ocean response ti the passage of
a mesoscale eddy off Bermuda, J. Geophys. Res., 104, 15537-15548.
Martin, J.M. & Milliman, J.D. (1997) EROS 2000 (European River Ocean
System). The western Mediterranean: an introduction. Deep-Sea Res. II,
44, 3-4, 521-529.
Marty, J.C., Chiaverini, J., Pizay, M.D. & Avril, B. (2002) Seasonal
and interannual dynamics of phytoplankton pigments in the Mediterranean Sea at
the France-JGOFS DYFAMED time-series station (1991-1999). Deep-Sea Res II, sous presse.
Marty, J.C., Nicolas, E., Miquel, J.C. & Fowler, S.W. (1994)
Particulate fluxes of organic compounds and their relationship to zooplankton
fecal pellets in the northwestern Mediterranean Sea. Mar. Chem., 46,
387-405.
Marty, J.C., Vescovali, I., Chiaverini, J. & Stock, A. (2000)
Temporal variability of hydrological conditions and related phytoplankton
pigment dynamics in the Mediterranean Sea: a nine year (91-99) study at DYFAMED
time-series station. 2nd JGOFS
Open Science Conference, Bergen (Norvège), 13-17 avril 2000, 132.
Mémery, L., Lévy, M., Vérant,
S. & Merlivat, L. (2002). The relevant time scales in estimating the
air-sea CO2 exchange in a mid-latitude region: a numerical study at the Dyfamed station (NW
Mediterranean Sea), Deep-Sea Res., sous presse.
Meybeck, M. (1982) Carbon, nitrogen and phosphorus
transport by world rivers. Amer. J. Sci., 282, 401-450.
Michaels, A F, Karl D M & D G Capone
(2001). Element stoichiometry, new production and nitrogen fixation. Oceanography, 14, 68-77.
Migon, C., Copin-Montégut,
G., Elégant, L. & Morelli, J. (1989) Etude de l'apport atmosphérique en
sels nutritifs au milieu côtier méditerranéen et implications biogéochimiques. Oceanol.
Acta, 12, 2, 187-191.
Migon, C. & Sandroni, V. (1999) Phosphorus in rainwater :
partitioning, inputs and impact on the surface coastal ocean. Limnol.
Oceanogr., 44, 4, 1160-1165.
Migon, C. & Sandroni, V. (2000) Fertilisation of surface seawater by
atmospheric inputs of phosphorus: investigations within the programme DYFAMED. 2nd JGOFS Open Science
Conference, Bergen (Norvège), 13-17 avril 2000.
Migon, C., Sandroni, V. & Béthoux, J.P. (2001) Atmospheric input of
anthropogenic phosphorus to the northwest Mediterranean during the oligotrophic
season. Mar. Environ. Res., 52, 413-426.
Migon, C., Sandroni, V., Marty, J.C., Gasser, B. & Miquel, J.C.
(2002) Transfer of atmospheric matter through the euphotic layer in the
northwestern Mediterranean: seasonal pattern and driving forces. Deep-Sea
Res. II, 49, 2125-2141.
Miquel, J.C., Fowler, S.W., La Rosa, J. & Buat-Ménard, P. (1994)
Dynamics of the downward flux of particles and carbon in the open northwestern
Mediterranean Sea. Deep-Sea Res. I, 41, 2, 243-261.
Montoya, J.P., Voss, M., Kähler, P. & Capone, D.G. (1996) A simple,
high precision, high sensitivity tracer assay for N2 fixation. Appl. Environ. Microb., 62, 986-993.
Moon van der Staay, S. Y., De Wachter, R. & Vaulot, D. (2001)
Oceanic 18S rDNA sequences from picoplankton reveal new eukaryotic lineages. Nature
409: 607-610.
Moran, X. A. G., Gasol, J. M., Pedros-Alio, C. & Estrada M. (2001)
Dissolved and particulate primary production and bacterial production in
offshore Antarctic waters during austral summer: coupled or uncoupled ? Mar.
Ecol. Prog. Ser., 222,25-39
Moutin, T. & Raimbault, P. (2002) Primary production, carbon export
and nutrient availability in western and eastern Mediterranean Sea in early
summer. J. Mar. Syst., sous presse.
Moutin, T., Raimbault, P., Golterman, H.L. & Coste, B. (1998) The
input of nutrients by the Rhône river into the Mediterranean Sea : recent
observations and comparison with earlier data. In : J.C. Amiard, B. Le
Rouzic, B. Berthet & G. Bertru (eds), Oceans, Rivers and Lakes : Energy and
Substance Transfers at Interfaces, Hydrobiologia, 373/374, 237-246.
Nelson, D.M., Tréguer, P., Brzezinski, M.A., Leynaert, A. &
Quéguiner, B. (1995) Production and dissolution of biogenic silica in the
ocean: revised global estimates, comparison with regional data and relationship
with biogenic sedimentation. Global Biogeochem. Cycles, 9, 359-372.
Obata H.,
Karatani H. & Nakayama E. (1993) Automated determination of iron in
seawater by chelating resin concentration and chemiluminescence, Anal. Chem., 65 :
1524-1528.
Olivier,
F. (2001) Etude de l’activité biologique et de la circulation océanique dans un
jet géostrophique : le front Almeria-Oran, PhD Thesis, Univ. Pierre et
Marie Curie, Paris.
Partensky, F., Blanchot, J., Lantoine, F., Neveux, J. & Marie, D.
(1996) Vertical structure of picophytoplankton at different trophic sites of
the subtropical northeastern Atlantic ocean. Deep-Sea Res. I, 43,
1191-1213
Raimbault
P., Diaz F., Boudjellal B. (1999b) Simultaneous determination of particulate
forms of carbon, nitrogen and phosphorus collected on filters using a
semi-automatic wet-oxidation procedure. Mar. Ecol. Progr. Ser., 180, 289-295.
Raimbault, P. & Pujo-Pay,
M. (1993) Activation d’absorption de nitrate par l’addition de phosphate dans
les eaux de l’Atlantique tropical. C.R. Acad. Sci., 316,
740-744.
Raimbault
P., Pouvesle W., Sempéré R., Diaz F., Garcia N. (1999a) A simple procedure for
simultaneous analysis of total and dissolved organic forms of carbon, nitrogen
and phosphorus in seawater using the wet-oxidation technic. Mar. Chem.,
66, 161-169.
Raimbault
P., Slawyk G., Boudjellal B., Coatanoan C., Conan P., Coste B., Garcia N.,
Moutin T., Pujo-Pay M. (1999c) Biomass, new production and export in the
equatorial Pacific at 150°W: Evidence for intense nitrogen recycling. J.
Geophys. Res. 104, 3341-3356.
Ridame
C. (2001) Rôle des apports atmosphériques d'origine continentale dans la
biogéochimie marine: Impact des apports sahariens sur la production primaire en
Méditerranée, Ph. D thesis, Univ. Paris VI, 246 pp.
Ridame C. & Guieu C. (2002) Saharan input of phosphorus to the
oligotrophic water of the open western Mediterranea. Limnol. Oceanogr., sous presse.
Sachs, J.P. & Repeta, D.J. (1999) Oligotrophy and nitrogen fixation
during eastern Mediterranean sapropel events. Science, 286, 2485-2488.
Sarthou, G. & Jeandel, C. (2001) Seasonal variations of iron
concentrations in the Ligurian Sea and iron budget in the western
Mediterranean. Mar. Chem., 74, 115-129.
Schöllhorn E. & Granéli E. (1993) Is the increase of flagellates in
coastal waters caused by changes in ratios of N, P and Si ? In: Toxic
Phytoplankton Blooms in the Sea, Smayda T.J. & Shimizu Y. (ed.). Elsevier
Science Publ., pp. 811-817.
Smayda T.J. (1990) Novel and nuisance phytoplankton blooms in the sea:
evidence for a global epidemic. In: Toxic Marine Phytoplankton, Granéli E.
(ed.). Elsevier Science Publ., pp. 29-40.
Steinberg, D.K., Carlson, C.A., Bates, N.R., Bates, R., Goldwaith, S.A.,
Madin, L.P. & Michaels, A.F. (2000) Zooplankton vertical migration and the
active transport of dissolved organic and inorganic carbon in the Sargasso Sea.
Deep-Sea Res. I, 47, 137-158.
Sterner, R. W. and Hessen, D. O., 1994. Algal nutrient limitation and
the nutrition of aquatic herbivores. Ann. Rev. Ecol. Syst., 25: 1-29.
Tanaka, T. & Rassoulzadegan, F. (2002) Full-depth profile (0-2000 m)
of bacteria, heterotrophic nanoflagellates and ciliates in the NW Mediterranean
Sea : Vertical partitioning of microbial trophic structures. Deep-Sea Res.
II, sous presse.
Thingstad, T.F., Zweifel, U.L., Rassoulzadegan, F. (1998) P limitation
of heterotrophic bacteria and phytoplankton in the northwestern Mediterranean. Limnol.
Oceanogr., 43, 88-94.
Thingstad, T.F., Hagström, A. & F. Rassoulzadegan (1997) Accumulation
of degradable DOC in surface waters : Is it caused by a malfunctioning
microbial loop ? Limnol. Oceanogr., 42, 2, 398-404.
Thingstad, T.F., Zweifel, U.L. & Rassoulzadegan, F. (1998) P
limitation of heterotrophic bacteria and phytoplankton in the northwest
Mediterranean. Limnol. Oceanogr., 43, 1, 88-94.
Thomson-Bulldis, A. & Karl, D.M. (1998) Application of a novel
method for phosphorus determinations in the oligotrophic North Pacific Ocean. Limnol.
Oceanogr., 43, 7, 1565-1577.
Touratier, F., Field, J.G. & Moloney, C.L. (2001) A stoichiometric
model relating growth substrate quality (C:N:P ratios) to N:P ratios in the
products of heterotrophic release and excretion. Ecol. Model., 139,
265-291.
Tranvik, C.J., Sherr, E.B. & Sherr, B.F. (1993) Uptake and
utilization of “ colloidal DOM ” by heterotrophic flagellates in
seawater. Mar. Ecol. Progr. Ser., 92, 301-309.
Touratier, F., Legendre, L. & Vézina,
A. (1999a) Model of bacterial growth influenced by
substrate C:N ratio and concentration. Aquat. Microb. Ecol., 19,
105-118.
Touratier, F., Legendre, L. & Vézina, A. (1999b) Model of
copepod growth influenced by the food carbon:nitrogen ratio and concentration,
under the hypothesis of strict homeostasis. J. Plankton Res., 21,
1111-1132.
Touratier, F., Legendre, L. & Vézina, A. (2000)
Northeast Water Polynya 1993: construction and modelling of a time series
representative of the summer anticyclonic gyre pelagic ecosystem. J. Mar. Syst., 27,
53-93.
Touratier, F., Field, J.G. & Moloney, C.L. (2001) A stoichiometric
model relating growth substrate quality (C:N:P ratios) to N:P ratios in the
products of heterotrophic release and excretion. Ecol. Model., 139,
265-291.
Tréguer P., Kamatani A., Gueneley S. & Quéguiner B. (1989) Kinetics
of dissolution of Antarctic diatom frustules and the biogeochemical cycle of
silicon in the Southern Ocean. Polar Biol., 9, 397-403.
Tréguer, P., Nelson, D.M., van Bennekom, A.J., Demaster, D.J., Leynaert,
A. & Quéguiner, B. (1995) The balance of silica in the world ocean: a re-estimate.
Science, 268: 375-379.
Tusseau-Vuillemin, M.H., Mortier, L. &
Herbaut, C. (1998) Modelling nitrate fluxes in an open coastal environment
(Gulf of Lions): transport versus biogeochemical processes, J. Geophys. Res.,
103, 4, 7693-7708.
UNEP (1996) The state of the marine and coastal environment in the
Mediterranean region, MAP Technical Report Series, 100, 142 pp.
Van Wambeke, F., Christaki, U., Giannakourou A.,
Moutin, T. & Souvemerzoglou, K. (2002) Longitudinal and vertical
trends of bacterial limitation by phosphorus and carbon in the Mediterranean
Sea. Microb. Ecol., sous presse.
Van Wambeke, F., Goutx, M., Striby, L., Sempéré, R., & Vidussi, F.
(2001) Bacterial dynamics during the transition from spring bloom to
oligotrophy in the Northwestern Mediterranean Sea. Relationships with
particulate detritus and dissolved organc matter. Mar. Ecol. Progr. Ser., sous presse.
Vidussi, F., Marty, J.C. & Chiaverini, J. (2000) Phytoplankton
pigment variations during the transition from spring bloom to oligotrophy in
the northwestern Mediterranean Sea. Deep-Sea Res. I, 47, 423-445.
Wells, M.L. & Goldberg, E.D. (1994) The distribution of colloids in
the north Atlantic and southern oceans. Limnol. Oceanogr., 39, 2,
286-302.
Zehr, J.P., Mellon, M.T. & Zani, S.
(1998) New nitrogen fixing microorganisms detected in oligotrophic oceans by
the amplification of nitrogenase (nifH) genes. Appl. Environ. Microbiol., 64, 3444-3450.
Zehr,
J.P., Waterbury, J.B., Turner, P.J., Montoya, J.P., Omoregie, E., Steward, G.F.,
Hansen, A. & Karl, D.M. (2001) Unicellular cyanobacteria fix N2 in the
subtropical North Pacific Ocean. Nature, 412, 635-638.




