click to go to ......
I. Quantification des réservoirs minéraux et organiques des éléments biogènes
I.1. Apports atmosphériques de nutriments/Marqueurs des sources d'émission
(2003 et 2004)
I.2.Stocks/flux d'éléments biogènes dans la colonne d’eau
II. Production/Biomasse/Processus d'assimilation
Estimation des flux particulaires et dissous associés à la production
II.1. Production
III. Fertilisations expérimentales des eaux de surface
IV.Modélisation
IV.1. Processus
IV.2. Evolution du système/Modèles prédictifs
Sorties mensuelles.
Les résultats acquis mensuellement, en
complément des données standard fournies par le SO DYFAMED, permettront
d’évaluer la variabilité temporelle de paramètres définissant
un état chimique et trophique donné.
Certains paramètres déjà acquis dans le
cadre du SO DYFAMED seront mesurés à nouveau lors de ces sorties
spécifiques, afin de prendre en compte leur variabilité à court
terme. Par exemple, des prélèvements pour dosage de pigments seront
effectués en concomitance avec les mesures de productions nouvelles
et régénérées.
| SORTIES MENSUELLES (2004 et 2005) |
|||
|
|
|
|
|
| Paramètres |
Personnes à bord |
Prélèvements pour
|
Effectué sorties
|
|
|
|
travaux à terre |
SO DYFAMED |
|
|
|
|
|
| Mise en oeuvre
CTD-Rosette |
1 |
|
|
|
|
|
|
|
| Stocks C,
N, P - Silice - |
|
|
|
| Absorption phosphate - |
1 |
oui |
|
| Productions nouvelle et régénérée |
|
|
|
|
|
|
|
|
| Picoplancton |
1 |
oui |
|
|
|
|
|
|
| Zooplancton |
2 |
|
|
|
|
|
|
|
| TEP - DOM |
1 |
oui |
|
|
|
|
|
|
| Production primaire (particulaire
+ |
|
|
|
| dissoute) - Enceintes - Filtrations
- |
|
oui (pigments) |
oui |
| Phytoplancton |
|
|
|
Sorties
saisonnières
En complément des sorties mensuelles, les trois
campagnes saisonnières prévues (janvier, mars et début septembre)
permettront de caractériser autant de configurations chimiques (disponibilité
des substances nutritives) et trophiques (production, stocks, processus
d’assimilation, flux) saisonnières typiques. Ces configurations
correspondent globalement aux trois périodes de mélange vertical,
de bloom et d’oligotrophie.
En cas d’indisponibilité d’un
navire de type Suroît, l’essentiel de ces travaux pourra être
effectué à l’aide de retours à terre, comme le permet la durée
du trajet Nice - site DYFAMED (3 heures).
I.
Quantification des réservoirs minéraux et organiques des éléments biogènes
I.1. Apports atmosphériques de nutriments/Marqueurs des sources d'émission (2003 et 2004)
Les flux atmosphériques de N,
P, Si, Fe seront quantifiés, et leur variabilité temporelle (événement/saison)
sera évaluée à travers la mesure i) du flux total (flux sec + flux humide),
source parfois significative de nutriments et de métaux pour les eaux de
surface, et ii) l'analyse de l'aérosol atmosphérique, qui permettra de
caractériser les diverses influences anthropiques et minérales et leur
variabilité. Le pas d'échantillonnage sera hebdomadaire pour l'aérosol et le
dépôt total, et les événements pluvieux seront systématiquement collectés, avec
le dépôt total ou spécifiquement.
La cinétique de solubilisation de la matière d'origine
atmosphérique, déterminante pour la disponibilité des substances nutritives,
sera étudiée expérimentalement. En particulier, une étude cinétique de
libération de P et Si sera menée sur des échantillons d’aérosol atmosphérique
(prélevés spécifiquement et sur le même pas de temps hebdomadaire) et sur des
particules de matériau-source caractéristique (e.g., poussières d’échappements
automobiles, cendres volantes, sol saharien).
Cette étude prendra en compte la dissolution en fonction de
l’éclairement : mesure comparative du taux de libération de P et Si sous
éclairement et dans l’obscurité. Il s’agira de choisir 40 échantillons
représentatifs de diverses situations météorologiques significatives sur la
centaine d’échantillons collectés au Cap Ferrat en deux ans et leur faire subir
une altération au contact d’une solution aqueuse avec des propriétés connues.
Cette solution pourra être soit acide, soit neutre/basique avec des complexants
pour simuler l’eau de mer. On étudiera sur ces échantillons la partition
soluble/insoluble des éléments nutritifs et du fer.
D’autres traceurs géochimiques (e.g., Al, Cr, Cu, K, Mn, Na,
Pb, S, V, Zn, etc.) seront analysés sur
les mêmes échantillons afin de déconvoluer les sources d'émission naturelles et
anthropiques de nutriments.
Dépôt total : 52
échantillons par an (2003-2004)
Aérosol : 52
échantillons par an (2003-2004)
Après
filtration du dépôt total, la fraction particulaire est pesée (estimation du
flux de poussières)
Conservation
des échantillons dissous à – 20 °C
C, N, P
particulaires (filtration d’aérosol) : colorimétrie automatique avec
oxydation humide (Raimbault et al., 1999b)
P, Si, Fe et
traceurs géochimiques (filtration d’aérosol) : fluorescence X
N (nitrate,
nitrite), P (orthophosphate), Si (silicate) dissous (dépôt total) :
colorimétrie automatique avec oxydation humide pour la détermination de C,
N et P totaux
Fe
dissous : ICP-AES
I.2.
Stocks/flux d'éléments
biogènes dans la colonne d’eau
La succession saisonnière des
niveaux de nutriments (N, P, Si) et de fer sera contrôlée dans les eaux de
surface et dans les pièges à particules (200 m). Pour les sels nutritifs, les
techniques d'analyse adaptées à des niveaux très bas en période de
stratification seront utilisées. Les flux de matière à 200 m seront disponibles
pour cette étude.
La concentration en orthophosphates sera mesurée par la méthode
MAGIC (MAGnesium Induced Coprecipitation) proposée par Karl & Tien (1992)
et modifiée par Thompson-Bulldis & Karl (1998). Les améliorations
méthodologiques apportées dans le cadre de BIOSOPE pourront être testées au cours
des campagnes MELISSA. Seront également dosés le phosphate organique dissous et
le phosphate organique particulaire (par fractions de taille 0,2, 0,6, 2 et 10
µm). Les améliorations récentes de la méthode classique de dosage au persulfate
permettent d'obtenir un niveau de précision de l'ordre de 1 nM, adaptée aux
milieux ultra-oligotrophes.
Des profils verticaux seront réalisés sur
l'ensemble de la colonne d'eau au site DYFAMED pour le suivi temporel de la
distribution des stocks dissous minéraux et organiques d'azote (nitrate,
nitrite et ammonium : NOD), du phosphore (phosphate : POD) et du COD. Les
techniques utilisées seront le dosage automatique par colorimétrie pour les
sels nutritifs et, pour la matière organique qui comprendra le COD, la méthode
semi-automatique décrite dans Raimbault et al. (1999a). A ces paramètres
s'ajouteront les réservoirs particulaires de carbone (COP), de l'azote (NOP) et
du phosphore (POP) par une méthode semi-automatique permettant le dosage
simultané de ces trois éléments (Raimbault et al., 1999b). L'ensemble de
ces mesures donnera accès aux rapports C:N:P dans la matière minérale et
organique.
De même, les stocks et les flux de Si seront mesurés, en
relation avec les études concernant l’azote, le phosphore et le fer (e.g.,
hypothèse de limitation de l’absorption de Si(OH)4 par la
disponibilité en fer, comme cela s’observe dans une zone HNLC australe). Les
fractions biogénique (BSi) et lithogénique (LSi) seront distinguées dans la
colonne d'eau sur 12 niveaux de profondeur.
Les concentrations de fer dissous seront
mesurées dans les 200 premiers mètres afin de suivre l’évolution des
concentrations dans la colonne d’eau en relation avec l’apport atmosphérique
(sorties mensuelles + sorties spécifiques, e.g. deux périodes contrastées dans
l'année; dans un premier temps les conditions oligotrophes, e.g. fin de l’été,
seront une priorité). Dans le cas des sorties spécifiques, des incubations in
vitro seront effectuées pour identifier les facteurs limitants et
déterminer le Ks du fer avec l’assemblage phytoplanctonique présent dans le
milieu pendant cette période. Ces expériences seront similaires à celles menées
dans le cadre des campagnes POMME (Blain et al., 2001); elles seront
conduites en microcosme et consistent à définir le rôle potentiel des éléments
nutritifs majeurs (N, P, Si), du fer et de poussières éoliennes sur la
production primaire. Ces expériences pourront être couplées à celles de la
diazotrophie afin de préciser le rôle du fer d’origine atmosphérique dans le
processus en Méditerranée.
Le matériel de prélèvement de la colonne d’eau, décrit dans
Guieu et al. (2002a), permet l’acheminement et la filtration en ligne de
l’eau prélevée dans les 200 premiers mètres de la colonne d’eau dans des
conditions ultra-propres. Le fer dissous sera dosé en flux continu par
chimiluminescence (Obata et al., 1993) pour la spéciation redox
(discrimination FeII/FeIII) et par polarographie pour la spéciation fer
réactif/fer complexé. Le broutage par le zooplancton pourrait être le processus
dominant du recyclage du fer biologique dans de nombreux environnements marins
(Hutchins et al., 1993). En effet, les conditions acides des intestins
des brouteurs facilitent le relargage du fer biologique des proies ingérées.
Dans l'Océan Austral, pendant la campagne de fertilisation SOIREE, il a été
montré que la régénération et l'assimilation du fer étaient du même ordre de
grandeur (Bowie et al., 2001). Qu'en est-il en Méditerranée ? Nous ne
savons pas sous quelle forme le fer est régénéré dans la phase dissoute (forme
inorganique Fe(II)/Fe(III) ou forme organique), ni si le fer régénéré est
assimilable ou plus facilement assimilable par le phytoplancton. La
caractérisation de la spéciation et de la biodisponibilité du fer régénéré
d'une part, et la quantification des processus de transfert entre les
différentes phases, d'autre part, seront réalisées grâce à des expériences de
broutage de communautés phytoplanctoniques naturelles marquées au fer-55, des
extractions sélectives liquide/solide et un comptage par scintillation liquide.
La variation saisonnière de ces processus sera étudiée lors des campagnes au
site DYFAMED aux différentes saisons.
a) C, N, P
► Campagnes
mensuelles (2004 et 2005)
L’évolution
temporelle des stocks de C, N et P sera mesurée.
Prélèvements :
rosette (12 échantillons : 10 L de surface ; 5 L 0-100 m ; 2 L
100-200 m ; 0,5 L 200-1000 m)
Empoisonnement
des échantillons par HgCl2 + réactifs supplémentaires pour NH4+
Analyse
colorimétrique automatique :
- Nitrate, nitrite, ammonium, phosphate,
silicate entre 0 et 1000 m
- C, N, P organiques particulaires entre 0
et 200 m
-
C, N, P
dissous entre 0 et 1000 m
► Campagnes
saisonnières (2004)
(comprend les
protocoles d’études d’impact des apports atmosphériques sur la production
primaire et l’assimilation d’azote – voir § II.1 et II.5.a)
Idem Campagnes
mensuelles, plus :
Les mesures
de C, N, P particulaires entre 0 et 200 m seront faites en distinguant les
différentes classes de taille (total ; < 10µm ; < 2 µm)
Manipulations
à bord : prélèvements ; filtration du matériel particulaire ;
dosage des sels nutritifs ; expérimentations et incubations 13C/15N ;
incubation en bacs sur le pont et sur une ligne in situ (24 h). Cette
stratégie pourra être répétée chaque jour avec des prélèvements en fin de nuit
(obtention de 5 profils et 5 séries d’expériences au cours de chaque sortie)
► Pièges à sédiments
fixes (2004 et 2005)
L’évolution
temporelle des rapports stoechiométriques C:N:P sera étudiée dans la matière
particulaire collectée dans les pièges à particules au site DYFAMED
(profondeurs : 200 et 1000 m)
Prélèvements :
une fraction de la matière collectée par godet (1 à 5 mg de matière)
Fréquence :
15 jours sur les deux pièges, soit 50 échantillons par an.
b) Silice
► Campagnes
mensuelles (2004 et 2005)
Mesure des
stocks BSi/LSi (colonne d’eau, 12 profondeurs + pièges à sédiment).
L’étude sera complétée par la mesure des taux de production de BSi ainsi que
des taux de dissolution à deux profondeurs (surface et base de la couche de
mélange).
► Campagnes
saisonnières (2004)
Mesures des flux et études de
processus (cinétiques d’absorption du Si, dissolution de BSi, cinétiques
d’absorption de l’acide silicilique et mesure des taux de régénération, effet
du fer sur les processus de co-limitation).
Analyse fine des populations
microphytoplanctoniques sur la base mensuelle du suivi saisonnier (prélèvements
en duplicats lugol-formol dans la couche mélangée) ainsi qu’au cours des trois
campagnes saisonnières. Ces travaux permettront en outre de tester les méthodes
d’évaluation de la biomasse siliceuse active par marquage fluorescent couplé à
l’analyse d’image. Ceci devrait améliorer notre connaissance de la dynamique
des populations de phytoplancton siliceux.
Les résultats attendus seront
mis en parallèle avec ceux acquis sur SOFI de façon à dessiner une première
estimation du cycle du silicium dans les écosystèmes méditerranéens.
Prélèvements : rosette
(1000 mL pour BSi et LSi, 1000 mL pour les vitesses de dissolution).
Profils complets (12
échantillons) pour BSi/LSi fractionné sur 2 et 10 µm, 2 échantillons pour les
vitesses de dissolution.
c) Cycle
biogéochimique du phosphore
► Campagnes
mensuelles (2004 et 2005)
Stocks
Mesure des
concentrations en orthophosphates par la méthode MAGIC (MAGnesium Induced
Coprecipitation, Karl et al., 1992) modifiée par Thomson-Bulldis &
Karl (1998).
Mesure du
phosphate organique dissous
Mesure du
phosphate organique particulaire (fractions de taille 0,2, 0,6, 2 et 10 µm).
Les adaptations récentes de la méthode au persulfate permettent d’obtenir une
précision de l’ordre de 1 nM adaptée aux milieux ultra-oligotrophes.
1 profil (12
profondeurs) pour l’ensemble des variables à l’exception du P particulaire par
fraction de taille qui ne sera mesuré qu’en surface et au maximum de
chlorophylle a.
Prélèvements :
300 mL x 2 (mesures MAGIC), 100 mL x 2 (P total), 500 mL (P particulaire)
Pas
d’embarquant spécifique : les analyses seront réalisées ultérieurement à
Marseille.
► Campagnes
saisonnières (2004)
En plus des
mesures de stocks ci-dessus, des mesures de flux seront réalisées à l’aide de 33P.
Flux
En parallèle
avec la mesure des taux d’absorption du phosphate par 33P, la
cinétique d’absorption des orthophosphates par les populations planctoniques
sera étudiée expérimentalement (avec une attention particulière pour la
compétition entre les hétérotrophes et les autotrophes de petite taille dans
les eaux oligotrophes méditerranéennes).
Le temps de
recyclage des orthophosphates sera étudié par marquage au 33P.
Plusieurs
profils (12 profondeurs) pour l’ensemble des variables et mesures de cinétique
d’assimilation de P par les organismes planctoniques en surface uniquement.
Prélèvements :
500 mL
1 embarquant
pour l’ensemble des mesures.
d) Fer dissous
► Campagnes
saisonnières (2004)
Prélèvements par pompage
propre in situ avec filtration à 0,2 µm : 250 mL par
profondeur entre 0 et 200 m (15 échantillons).
Analyses à terre
(chimiluminescence).
Expériences de
fertilisation à bord
Prélèvement d’eau :
profondeur en fonction de la stratification de la colonne d’eau, de toute façon
< 50 m.
38 bouteilles de 4 L.
L’incubation s’étend sur toute la durée du temps sur site.
Analyses à terre.
Volume d’eau nécessaire : 12 L (surface ; 6 échantillons)
e)
Quantification de la spéciation et de la biodisponibilité du fer régénéré par
le broutage du zooplancton
► Campagnes
saisonnières (2004)
La spéciation et la
biodisponibilité du fer régénéré seront quantifiées par des techniques
isotopiques, avec l’utilisation du radiotraceur 55Fe et d’un
comptage à scintillation liquide. Les expériences seront adaptées de Bowie et
al. (2001, voir figure 4) qui ont étudié les taux de
régénération du fer par le broutage du micro- et du méso-zooplancton nourris
avec i) des bactéries et du "petit" phytoplancton (< 2 µm) et ii)
du "grand" phytoplancton (> 2 µm).
Trois jours avant les
expériences de broutage, le plancton de classe de taille < 2 µm et 2-20 µm
est incubé avec du 55Fe à lumière et température ambiantes. Après
trois jours d’incubation, les cellules sont collectées sur des filtres en
polycarbonate de porosité 2 µm et 20 µm et rincées avec une solution de
Ti(III)EDTA-citrate pour éliminer le fer extracellulaire (Hudson & Morel,
1989 ).
Pour les expériences de
broutage par le micro-zooplancton, les cellules (de tailles < 2 µm) marquées
au 55Fe sont filtrées puis remises en suspension dans 1 L d’eau de
mer. Après 24 h, la spéciation du 55Fe (Fe organique et Fe
inorganique Fe(II)/Fe(III)) dans la phase dissoute est mesurée par extraction
sélective sur résine (C18 et 8-hydroxyquinoleine ; Obata et al.,
1993 ; Elbaz-Poulichet et al., 1994). L’activité du 55Fe
est également mesurée dans les fractions >20 µm, 2-20 µm et 0,2-2 µm
(rincées avec une solution de Ti(III)EDTA-citrate ; Hudson & Morel,
1989).
Les expériences destinées à l’étude de la régénération par le
broutage du mesozooplancton sont similaires. Les cellules (> 2 µm) marquées
au 55Fe sont remises en solution dans 1 L d’eau de mer et des
copépodes sont ajoutés (4 L-1). La spéciation et l’activité du 55Fe
dans les fractions > 20 µm, 2-20 µm, et 0,2-2 µm sont également mesurées.
Les contrôles consistent à remettre en suspension les cellules
marquées dans de l’eau de mer filtrée à 0,2 µm.
Pour chaque expérience, la biodisponibilité du fer régénéré est
quantifiée par des expériences de croissance de cellules non marquées dont le
milieu de culture est la phase dissoute ayant subie le broutage.
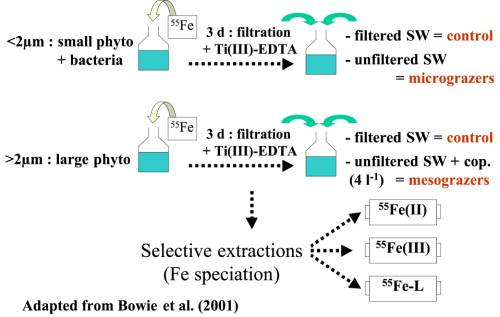 |
Figure 4:
Chemical speciation and bioavailability of iron regenerated by zooplankton
grazing (adapted from Bowie et al., 2001).
II. Production/Biomasse/Processus d'assimilation
Estimation des flux particulaires et dissous associés à la
production
Le projet
utilisera les mesures mensuelles standard de production primaire particulaire
acquises dans le cadre du SO DYFAMED ainsi que les mesures effectuées lors des
campagnes saisonnières spécifiques (3 campagnes en 2004). Avec le projet sera
développé un protocole de mesure de la production dissoute aussi bien pour les
mesures mensuelles que pour les mesures des campagnes saisonnières. Le taux
d'assimilation du carbone (production primaire) sera mesuré par marquage au 14C.
En outre, lors des expérimentations à l’azote-15, la production primaire sera
spécifiquement estimée par marquage au 13C lors des campagnes
MELISSA sur 6 niveaux de profondeur (technique du double marquage 13C-15N).
La production nouvelle sera quantifiée par
la mesure d'absorption de nitrate (15N-NO3) et de la
fixation d’azote (15N -N2).
La production régénérée sera quantifiée
par la mesure d’absorption d’ammonium (15N-NH4). Ces
mesures de production seront complétées par des estimations des flux de
régénération : régénération d'ammonium, nitrification (oxydation de
l'ammonium), d'excrétion d'azote organique dissous (NOD), selon les protocoles
décrits dans Raimbault et al. (1999c), pour déterminer le devenir
immédiat de la matière photosynthétisée et de la fraction potentiellement
exportée.
Les activités bactériennes seront suivies
dans la colonne d’eau de surface (0-200 m) : production et phosphatase
alcaline par classe de taille.
En parallèle, la fraction exportée sera
évaluée à l’aide de pièges dérivants (suivi sur 48 h, pas de temps de 6 h),
dans le cadre des campagnes saisonnières.
a)
Production primaire particulaire et dissoute
► Campagnes
mensuelles (2004 et 2005)
Production
primaire particulaire et dissoute : prélèvements (10 mL) par enceintes LET
GO à 10 profondeurs
Filtrations
à bord : 1 m2 de paillasse ; 1 personne
Mesure
de la production dissoute : protocoles de Karl et al. (1998) et
Moran et al. (2001)
Autres
manipulations au retour à terre (conservation des échantillons à 4°C)
Nombre
d’échantillons par jour : 10 (production primaire)
► Campagnes
saisonnières (2004)
Idem « Campagnes
mensuelles », plus :
2
personnes à bord, 3 m2 de paillasse (comprend les mesures de
pigments, voir § II.2.a, et les expériences de fertilisation, voir § III)
b)
Production nouvelle/Production régénérée
►
Campagnes mensuelles (2004 et 2005) et saisonnières (2004)
Utilisation
de traceurs isotopiques
Production
primaire : marquage au 13C dans expérimentation à l’azote-15
Production
nouvelle (assimilation de nitrate et fixation d‘azote moléculaire) : 15N
Production
régénérée : 15N
Régénération
de l’ammonium : 15N
Nitrification :
15N
Excrétion
d’azote organique dissous : 15N
c) Activité
bactérienne hétérotrophe
Les
activités bactériennes seront suivies dans la colonne d’eau de surface (0-200
m) : production et phosphatase alcaline par classe de taille, expériences
d’enrichissement à différentes profondeurs sur la verticale. Ces mesures ont
pour but :
- de mesurer la production bactérienne et
l’activité phosphatase alcaline in situ, en relation avec les cycles du
phosphore et du carbone.
- de suivre la variabilité des facteurs
limitant les bactéries hétérotrophes sur la colonne d’eau (zone euphotique
essentiellement, approximativement les 150 premiers mètres), en fonction de
certaines situations saisonnières types, en réalisant des expériences
d’enrichissement en azote, phosphore minéral et glucose.
►
Campagnes saisonnières (2004)
1 profil vertical
à 12 niveaux (épifluorescence, cinétique de l’activité phosphatase globale,
détection de l’activité phosphatase au niveau cellulaire, production
bactérienne classe de taille 0,2 et 0,6 µm).
Expérience
d’enrichissement avec suivi de production bactérienne et d’activité
phosphatase.
1 personne à bord
Surface de
travail : 1,5 m dans le container radioactif + 1,5 m en dehors du
container.
II.2.
Biomasse
phytoplanctonique et diversité génétique du picophytoplancton
Les variations
saisonnières de la biomasse phytoplanctonique totale et spécifique
(chlorophylle a + pigments spécifiques, analysés par HPLC) seront
étudiées à partir de mesures mensuelles standard (acquises dans le cadre du SO
DYFAMED) et au cours des campagnes tri-annuelles spécifiques au projet.
Une analyse fine des populations microphytoplanctoniques (base
mensuelle du suivi saisonnier + trois expériences saisonnières) est proposée.
Ces actions pourraient aussi servir à tester les méthodes d’évaluation de la
biomasse siliceuse active par marquage fluorescent couplé à l’analyse d’image.
Résultats attendus sur la dynamique des populations de phytoplancton siliceux.
Les résultats seront mis en parallèle avec ceux acquis en 2000 à la station
SOFI (talus continental au large de Marseille, conditions mésotrophes) où des
mesures de flux d’absorption de Si(OH)4 ont été réalisées par
marquage au 32Si, ainsi que des mesures de stocks et de flux
verticaux de silice biogénique et lithogénique.
Les variations saisonnières de l'abondance des populations du
picophytoplancton seront étudiées par cytométrie en flux (Prochlorococcus,
Synechococcus, picoeucaryotes +/- virus).
L’évolution saisonnière de la diversité phylogénétique du
picophytoplancton sera étudiée à 4 à 6 profondeurs par des méthodes
moléculaires : le picoplacton eucaryote sera étudié par analyse du gène de
l’ARNr 18S en DGGE (denaturing gradient gel electrophoresis) et séquençage des
bandes d’intérêt (e.g. correspondant à des génotypes abondants, ubiquistes ou
présents toute l’année, etc.) et/ou PCR (Polymerase Chain Reaction)
quantitative. Les prélèvements seront réalisés sur les mêmes profils que les
mesures HPLC/Pigments. Ces données devraient permettre d’estimer l’abondance
relative des principaux groupes du picoplancton eucaryote (Prasinophyceae,
Pelagophyceae, Prymnesiophyceae).
Prochlorococcus sera étudié par analyse du gène
fonctionnel pcb codant pour le principal complexe collecteur de photons
par la méthode de PCR-RFLP (Random Fragment Length Polymorphism). Elle consiste
à amplifier puis digérer ce gène par l’enzyme de restriction Haelll afin
d’obtenir des profils de restriction sur gel d’électrophorèse (Metaphor).
L’étude des variations temporelles de ces profils permettra notamment de suivre
la mise en place de la stratification verticale des populations de Prochlorococcus
après le mélange hivernal.
Détection de la présence du gène nifH.
La présence dans la colonne d’eau du gène nifH,
indicateur d’organismes potentiellement fixateurs d’azote (comme les
cyanobactéries Trichodesmium ou certaines espèces de Synechocystis)
sera également étudié à 1 ou 2 profondeurs au cours du suivi annuel en
utilisant des amorces spécifiques de ce gène et en essayant de l’amplifier
sélectivement par PCR. Les échantillons dans lesquels un produit de PCR sera
détecté seront étudiés plus avant en séquençant le produit obtenu afin de
récolter une information phylogénétique sur le ou les organismes d’où provient
ce gène. Ces données permettront de mettre en évidence un éventuel cycle annuel
des organismes fixateurs d’azote à la station DYFAMED.
Les paramètres ainsi mesurés sont :
- Concentration du picoplancton (bactéries, Prochlorococcus,
Synechococcus, picoeucaryotes)
- Diversité des échantillons naturels de picoeucaryotes
visualisée par un profil de bandes sur gel d’électrophorèse (DGGE)
- Séquences du gène de l’ARN 18S et arbres phylogénétiques
- Abondance relative des divers groupes du picoplancton
eucaryote
- Diversité génotypique de Prochlorococcus après amplification
et digestion du gène pcb
- Séquences du gène nifH
Pour la biomasse zooplanctonique, les
migrations nycthémérales sont prises en compte avec un échantillonnage
jour/nuit sur un cycle d'au moins 48h, avec pour chaque point d'échantillonnage
:
1) un trait
vertical 0-200m avec un filet triple 200µm pour estimation de la biomasse
sestonique et de la taxonomie (paramètres standards internationaux);
2) un trait
double-oblique sur 0-200m avec un filet de vide de maille 200µm afin de
récolter les organismes vivants à mettre en expérience dans des conditions
nutritionnelles naturelles ou limitatives, afin : i) d'estimer l'activité et la
sélectivité alimentaire (mesures pigmentaires et incubations); ii) de mesurer
la fécondité des populations; iii) d'évaluer les processus d'égestion.
Ces
expériences seront conduites sur les espèces dominantes.
a)
Phytoplancton
►
Campagnes mensuelles (2004 et 2005)
Voir les
mesures standard de pigments effectuées dans le cadre du SO DYFAMED.
Echantillons de
pièges à sédiments fixes (200 et 1000 m)
►
Campagnes saisonnières (2004)
Description de
la distribution verticale des groupes phytoplanctoniques spécifiques par la
mesure des pigments par HPLC, en relation avec la distribution détaillée des
divers nutriments. Détermination de la variabilité temporelle de ces paramètres
sur 72 h. La distribution sera détaillée au niveau des différentes nutriclines
(12 profondeurs ou plus). Suivi des évolutions temporelles (sur 48 h) des
populations phytoplanctoniques dans des enceintes dopées par un nutritifs
susceptible d’être apporté par voie atmosphérique.
Mesure
de pigments sur les échantillons de pièges dérivants de type PPS 5 (1 aliquote,
soit 1/8ème)
Prélèvements :
CTD rosette (pigments) à 12 profondeurs
Nombre d’
échantillons par jour : 24 (pigments) + enceintes (cf § III)
2 profils par
24 h (jour + nuit) pour les pigments (4-5 jours)
1 profil
journalier de production primaire particulaire et dissoute (4 jours)
Dosage des
pigments (caroténoïdes et phaeopigments) sur le matériel des pièges dérivants
(4 échantillons par sortie)
b)
Picophytoplancton
►
Campagnes mensuelles (2004 et 2005)
1)
Cytométrie : 2 mL
fixés au glutaraldéhyde 0,2% (15 mn), congélation dans l’azote liquide puis
stockage à – 80°C. Prélèvements à toutes profondeurs. 2 échantillons par
profondeur.
2)
Diversité et abondance des picoeucaryotes : 1,5 à 3 L (selon la biomasse chlorophyllienne)
préfiltrés sur 3 µm et récoltés sur filtre Supor 0,45 µm (filtres en cascade),
congelé dans un tampon ADN. Prélèvements à 3 profondeurs en période homogène,
davantage (4 à 6) dès le début de la stratification de la colonne d’eau (e.g.,
5 m, nitracline, maximum de chlorophylle, 20 m au-dessus du maximum de
chlorophylle). 1 échantillon par profondeur (2004 seulement)
3)
Diversité de Prochlorococcus : 1 L filtré sur Supor 0,2 µm et congelé à sec dans un
cryotube. Profondeurs/nombre d’échantillons : idem (2) (2004 seulement)
►
Campagnes saisonnières (2004)
Idem
« Campagnes mensuelles », plus analyses cytométriques et analyses de
la diversité des picoeucaryotes par DGGE à T0 et en fin d’incubation
des différents échantillons incubés à bord après ajout de traces de différents
éléments.
Remarque : la participation aux campagnes
saisonnières sur ce poste se limitera à une sortie (mars ou septembre).
c) Zooplancton
Paramètres mesurés :
taux d’ingestion des espèces dominantes par groupe de proies potentielles, y
compris les proies hétérotrophes ; sélectivité ; production gonadique
des espèces dominantes ; taux d’égestion des espèces dominantes.
► Campagnes
mensuelles (2004 et 2005)
Prélèvements par filets WPII
de 200 µm horizontaux et obliques ; bouteilles Niskin
Volume d’eau prélevé :
60 L ; profondeur : 0-200 m ; 12 prélèvements / 24 h ; 44
échantillons issus de l’incubation de certains de ces prélèvements / 24 h
Manipulations à bord :
tri des prélèvements à la loupe binoculaire, mesure de la fluorescence
intestinale et incubations pour 1) mesures de broutage 2) mesures de la
production d’œufs et 3) mesure de la production de pelotes fécales
2 personnes à bord
2 m linéaires de paillasse
II.3. Flux particulaires et
dissous
Les données régulières de
flux à 200 m acquises dans le cadre du SO DYFAMED permettront d’appréhender les
variations temporelles et verticales des flux particulaires en fonction des
différentes configurations de limitation chimique.
Voir aussi § II.1
► Campagnes
mensuelles (2004 et 2005)
a) Distribution saisonnière
des TEP dans la colonne d’eau - Etude de la production et de la colonisation
bactérienne des TEP en relation avec le statut trophique du milieu
Pour savoir si les TEP
constituent ou non un support privilégié pour le développement des bactéries,
la colonisation bactérienne sera étudiée ainsi que le nombre de bactéries
actives fixées à ces particules prélevées à différentes profondeurs. Cette
étude in situ complétera les études en microcosmes en cours
(interactions TEP-bactéries et interactions TEP-organismes filtreurs, en
l’occurrence des copépodes). Les paramètres mesurés sont : l’abondance et
le spectre de taille des TEP, l’abondance et la caractérisation des bactéries
associées aux TEP.
Prélèvements de 200 mL à 10,
30, 50, 90, 130, 200, 500, 1000 et 2000 m.
Pas de personne
spécifiquement embarquées pour ce travail. Manipulations à terre (conservation
des échantillons à T ambiante si retour < 5 heures, sinon congélation à –
20°C).
b) Variations
saisonnières des propriétés d’adhésion des TEP - Caractérisation microscopique
des TEP à partir d’échantillons purifiés et concentrés de TEP provenant de la couche euphotique
Les objectifs sont : i)
déterminer les rôles de la turbulence et de la dégradation bactérienne dans les
processus d’agrégation/désagrégation, afin de prédire les variations dans les
flux particulaires en fonction de la profondeur et de la saison (limitation
saisonnière) ; ii) caractériser chimiquement les TEP et révéler les
variations saisonnières de leur composition liées aux conditions de limitation
en nutriments.
Prélèvements par bouteilles
Niskin de 10 L à 25, 50, 100, 250, 500 et 1500 m pour les expériences de
coagulation
Prélèvement de 10 L à 25 m
pour purification et concentration des TEP.
Manipulations à bord :
Préfiltration des échantillons ; purification et concentration des TEP pour
caractérisation microscopique et analytique.
2 m2 de paillasse
requis.
► Campagnes
saisonnières (2004)
a) Abondance et spectre
de taille des TEP dans la colonne d’eau (complément aux études mensuelles) -
Colonisation bactérienne des TEP à court terme en relation avec le statut
trophique du milieu
Protocoles : idem
sorties mensuelles a) + prélèvement (2 L) toutes les 4 heures à 2 profondeurs.
1 personne à bord, 60 cm de
paillasse.
b) Caractérisation
chimique de la HWM DOM
La nature et l’abondance de
ce matériel semi-labile recyclé aux échelles mensuelle et annuelle seront
déterminées par la caractérisation de la fraction de haut poids moléculaire
(HWM, environ 30% de la DOM totale). La fraction de HWM de la DOM sera
récupérée (campagnes saisonnières) par ultrafiltration à bord. La spectroscopie
RMN et les techniques électrophorétiques seront utilisées pour étudier la
distribution des groupes organiques fonctionnels dans les isolats de HWM DOM. Différents
monomères organiques (sucres, aminoacides, lipides) seront isolés et
caractérisés par GC, HPLC et GC-MS.
En complément, des
expériences in situ et en culture permettront de mesurer la consommation
de biomolécules dissoutes et la production bactérienne. Ceci contribuera à
évaluer la disponibilité de tels substrats naturels vis-à-vis du
bactérioplancton. Plus globalement, cette approche permettra d'expliciter les
paramètres qui contrôlent l'occurrence de la DOM sous différentes dynamiques
nutritives et trophiques.
c) Relation entre les
stocks de HWM DOM et de TEP
Afin d'étudier l'hypothèse de
formation des TEP par coagulation des AcylPolySaccharides (APS), la composition
chimique des HMW DOM et des TEP sera analysée, à travers les données
saisonnières acquises au site DYFAMED et des expérimentations en culture. Les
TEP seront purifiées et concentrées selon la méthode magnétique (Mari &
Dam, en prép.) également utilisée pour caractériser chimiquement les TEP
(campagnes mensuelles). Si cette hypothèse est vérifiée, une fraction
importante de la DOM peut être exportée via la formation de TEP, ce qui
souligne le rôle que jouent les TEP entre les cycles de DOM et de matière
organique particulaire.
d) Propriétés
d’adhésion et caractérisation chimique des TEP
Les macromolécules composant
la matière organique colloïdale (30 à 40% du pool de DOM ; Benner et al.,
1992 ; Santschi et al., 1995) sont supposées être à l’origine de la
formation des TEP. Mais si l’agrégation de la HWM DOM en neige marine par
l’intermédiaire des TEP est une voie majeure d’excrétion de la DOM de la
surface des océans, ce processus est peu connu. La nature chimique des TEP et
leurs propriétés d’adhésion seront étudiées en relation avec la disponibilité
saisonnière des nutriments et en fonction de la profondeur. Les propriétés
d’adhésion des TEP seront étudiées au laboratoire (expériences de coagulation
en conditions contrôlées sur 4 niveaux de turbulence, un dispositif oscillant
produisant les différents niveaux de turbulence). Un exercice de modélisation
permettra de déterminer le coefficient d’adhésion des TEP.
Remarque : Les analyses qui requièrent un
matériel indisponible à Villefranche (RMN, GC-MS) seront effectuées sous la
responsabilité de Dan Repeta.
Prélèvements en
bouteilles Niskin :
- 10 L à 25, 50,
100, 250, 500 et 1500 m pour les TEP (purification et concentration) et 200 L à
25, 500 et 1500 m pour HMW DOM (purification et concentration) ;
- 200 L à 25 m
pour expériences sur la composition chimique des TEP et leurs propriétés
d’ adhésion en fonction de l’activité bactérienne (15 jours d’incubation)
Manipulations à
bord : 2 personnes, 3 m2 de paillasse
Collaborations :
Adrian B. Burd (Univ. of Athens, Georgie, USA)
Peter H. Santschi (Univ. of Texas A&M, Galveston,
Texas, USA)
Daniel J. Repeta (Woods Hole Oceanographic Institute,
Woods Hole, USA)
II.5.
Processus d'assimilation
a) Fixation
biologique de N2
Si dans certaines
zones océaniques, P est l'élément limitant, c'est d'abord parce que N ne l'est
pas. Qu'en est-il en Méditerranée ? La fixation d'azote atmosphérique a-t-elle
lieu, et dans ce cas, quelle est son intensité ? Jusqu'à présent, la
quantification de la diazotrophie en Méditerranée a été indirecte (e.g., bilans
à l'échelle d'un bassin). Ce processus doit à présent être quantifié
directement. Deux approches sont prévues ici :
1) Utilisation de l'isotope stable 15N,
d'après le protocole décrit par Montoya et al. (1996), parallèlement aux
mesures classiques des flux d'autres espèces chimiques d'azote (nitrate et
ammonium). Les mesures de diazotrophie doivent être effectuées séparément sur
la fraction picoplanctonique (bien que l’analyse de génomes complets de Prochlorococcus
et Synechococcus semble indiquer qu’aucun de ces deux organismes ne
possède le gène nifH et donc la capacité de fixer l’azote) et la
fraction nanoplanctonique (taille > 3 µm) qui, malgré sa faible abondance
dans les eaux oligotrophes (5 à 30% de la chlorophylle totale), pourrait être
le siège principal de la diazotrophie (Zehr et al., 2001).
2) L'approche génétique permettra
d'identifier dans les échantillons naturels (clonage et séquençage) les espèces
possédant le gène nifH, c'est-à-dire potentiellement capables de fixer N2.
Malgré les nombreuses publications suggérant la fixation de N2
atmosphérique en Méditerranée, aucune vérification directe n'a été faite à ce
jour. Une première étape consistera à
rechercher la présence du gène nifH à une ou deux profondeurs au cours
du suivi saisonnier. Cette étude qualitative et les mesures quantitatives de 15N
sont complémentaires.
Il faut ajouter
que d'une part, la fixation d'azote ne peut se faire en l'absence de tout autre
substance nutritive, et l'étude du processus de diazotrophie se fera en
connaissance des autres apports nutritifs, notamment le phosphate et le fer.
Ceci sera particulièrement important en conditions d'oligotrophie, quand les
apports atmosphériques constituent la seule source possible d'alimentation pour
les eaux stratifiées du large. D'autre part, l'assimilation d'azote résultant
du processus de diazotrophie doit conduire à des rapports cellulaires N/P
supérieurs à 16 chez les organismes concernés. Il sera donc important de suivre
l'évolution des rapports C/N/P dans la fraction particulaire, autant que
possible dans la fraction contenant préférentiellement les fixateurs d'azote
(nano- et picoplancton), notamment dans les configurations saisonnières
favorables à leur développement.
b) Limitation par P et assimilation
compétitive
Quelle est la
relation entre la limitation par les nutriments et la diversité des populations
microbiennes ? Cet aspect sera abordé par des expériences in vitro :
l'eau de surface du site DYFAMED sera utilisée pour dopage/incubation au laboratoire.
Ajouts de NH4+, PO43- et glucose
(en tant que source de carbone). L'effet de l'enrichissement sera examiné à
intervalles réguliers. Cette approche est très complémentaire des travaux de
dopage/incubation proposés en milieu naturel. Les paramètres de contrôle seront
ici l'incorporation de leucine, de 14C, les biomasses bactérienne et
phytoplanctonique, l'activité bactérienne, la cinétique d'assimilation et le
temps de turnover (marquage par 33P) et les mesures de P soluble
réactif, POD, POP, COD et COP. Des échantillons d'eau seront prélevés dans la
colonne d'eau et incubés à température in situ et la cinétique de
consommation d'oxygène sera mesurée.
c)
Métabolisme bactérien
L'efficacité
de croissance bactérienne sera estimée dans la couche de surface (0-200 m) en
mesurant la l’abondance, la production et la respiration bactériennes. Un cycle
d'un an pourrait débuter à l'été 2003 dans le cadre d'une comparaison avec des
systèmes côtiers : point B de la baie de Villefranche-sur-mer, baie de
Palma ainsi que l'estuaire et le panache de l'Escaut (travail post-doctoral de
N. Gonzalez-Benitez, LOV Villefranche).
Le métabolisme hétérotrophique dans la colonne d’eau sera
examiné en relation avec les flux de COP et de DOC (processus de transport
vertical et de régénération). Les bactéries sont-elles des producteurs ou des
reminéralisateurs ? L’objectif est de quantifier la contribution de la
respiration planctonique aux processus de reminéralisation dans la colonne
d’eau, en particulier dans la couche aphotique. Selon des travaux récents sur
la colonne 0-2000 m au site DYFAMED (Tanaka & Rassoulzadegan, 2002), les
profils verticaux d’abondance et de biomasse microbiennes dans la colonne d’eau
montrent que le modèle log-log de décroissance en fonction de la profondeur ne
se vérifie pas dans la couche mésopélagique. Cela suggère des mécanismes de
transport de la matière organique différents des couches mésopélagique à
bathypélagique.
III. Fertilisations expérimentales des eaux de surface
Les échantillons de pluie
collectés au Cap Ferrat ont déjà permis de calculer l'impact théorique d'un
apport atmosphérique significatif en phosphore sur la dynamique
phytoplanctonique en conditions oligotrophes : par exemple, un flux de 17 µmol
P réactif m-2, observé en période estivale, pourrait induire, en
admettant un rapport C/P de 106, une production de l'ordre de 0,02 g C m-2
(Migon & Sandroni, 1999). La vérification expérimentale d'un tel impact
sera effectuée in situ : des enceintes transparentes de type LET GO (4,5
litres), enrichies avec une quantité de phosphore (ou de tout autre élément
intervenant dans la limitation de la production, comme l’indiqueront les études
de caractérisation saisonnières) correspondant à un tel apport compte tenu d'un
effet de dilution sur une colonne d'eau typique de 5 m, seront mouillées au
site DYFAMED. Après incubation, l'évolution du milieu sera observée :
concentrations en sels nutritifs, production primaire, biomasses
phytoplanctoniques totale et spécifique (mesures pigmentaires par HPLC),
abondance et dynamique des populations du picoplancton, etc., permettant ainsi
d'appréhender les paramètres chimiques qui régulent la production primaire, la
cinétique d'assimilation de N, P et Si par les diverses espèces, ou encore
l'aptitude d'une espèce donnée à fixer le carbone du milieu. A noter que cette
approche, qui sera utilement complétée par des expériences en cultures, a déjà
révélé une réaction rapide (dès 24 h d’incubation) de la plupart des classes
phytoplanctoniques (Chambard, 1999; Magro et al., 2000; Migon &
Sandroni, 2000).
Par ailleurs, l'influence de la disponibilité en P ou Fe sur le
cycle de N et sur la production primaire sera examinée au moins une fois par
campagne par des expériences d'enrichissement (Fe + phosphate) réalisées en
bacs sur des échantillons de surface avec des incubations de l'ordre de 48 h.
Dans le même ordre d'idées, l'impact d'une co-limitation en fer et en phosphore
sera étudiée expérimentalement (en particulier, l'hypothèse de la limitation de
l'absorption de Si(OH)4 par la disponibilité en fer, comme cela a
déjà été démontré en zone HNLC australe). La problématique d'enrichissement
chimique du compartiment pélagique sera étroitement couplée aux études de
dissolution du matériel atmosphérique dans la couche marine de surface. Le rôle
des poussières minérales (dissolution de la silice lithogénique) sera évalué.
Quelle est la relation entre limitation
par les nutriments et diversité des populations microbiennes ? Cet aspect sera
abordé par des expériences in vitro : l’eau de surface du site DYFAMED
sera utilisée pour dopage/incubation au laboratoire. Ajouts de NH4+,
PO43- et glucose (en tant que source de carbone). L’effet
de l'enrichissement sera examiné à intervalles réguliers. Cette approche est
très complémentaire des travaux de dopage/incubation proposés en milieu
naturel. Les paramètres de contrôle seront ici l’incorporation de leucine, de 14C,
les biomasses bactérienne et phytoplanctonique, l’activité bactérienne, la
cinétique d’assimilation et le temps de turnover (marquage par 33P)
et les mesures de P soluble réactif, POD, POP, COD et COP. Des échantillons
d’eau seront prélevés dans la colonne d’eau et incubés à température in situ
et la cinétique de consommation d’oxygène sera mesurée.
Les algues comme les bactéries
hétérotrophes sont capables de développer l’expression d’une activité
ectoenzymatique, la phosphatase alcaline, qui permet d’obtenir du phosphate
sous forme minérale après hydrolyse de phosphore organique dissous. Le suivi
simultané de l’activité phosphatase et de la production bactérienne in situ,
ainsi que des expériences d’enrichissement (in vitro) en N, P minéral,
glucose, seuls et/ou en combinaison, ont permis d’établir un gradient
horizontal de limitation dans les eaux de surface en Méditerranée, mais aussi
un gradient de limitation sur la
verticale dans la zone euphotique (Van Wambeke et al., 2002). A la
station DYFAMED, en septembre, les bactéries hétérotrophes passent d’un stade
de limitation en phosphate à un stade de limitation en carbone labile à une
profondeur où le phosphate est toujours indétectable, et qui correspond à un
temps de turnover de la phosphatase alcaline d’environ 100 h (Van Wambeke et
al., 2002). Des expériences d’enrichissement seront menées à différentes
profondeurs sur la verticale avec pour objectif : i) de mesurer la production
bactérienne et l’activité phosphatase alcaline in situ, en relation avec
le cycle du phosphore, et du carbone; ii) de suivre la variabilité des facteurs
limitant les bactéries hétérotrophes sur la colonne d’eau (zone euphotique
essentiellement, les 150 premiers mètres environ), en fonction de certaines
situations saisonnières types, en réalisant des expériences d’enrichissement en
N, P minéral, et glucose (voir § II.1.c).
► Campagnes saisonnières
(2004)
Protocoles :
1-2
expérience(s) en enceinte LET GO de 4,5 L : effets d’un apport nutritif
sur la production et sur la composition phytoplanctonique spécifique (pigments)
Enceintes à 3
et 10 m (couche mélangée et surface, selon profil)
IV.
Modélisation
L'approche
modélisatrice sera intégrée dès sa mise en route du projet. Plusieurs échelles seront couvertes, allant
du processus à la variabilité interannuelle.
Jusqu’à présent, les
modèles 1-D appliqués au site DYFAMED ne prenaient en compte que la limitation
par N (Andersen & Nival, 1988; Lévy et al., 1998a; Tusseau et al.,
1998), et dernièrement la limitation par Si (Olivier, 2001). Cependant, il
n'existe pas actuellement de modèle biogéochimique pour la Méditerranée faisant
intervenir le P, ni les apports atmosphériques en sels nutritifs, les processus
de dissolution, etc. Les questions scientifiques importantes qui motivent les
modélisateurs et qui seront couvertes par un programme d'envergure au site
DYFAMED sont :
- la co-limitation de
la photosynthèse, avec un aspect fondamental et particulièrement bien adapté à
la Méditerranée, à savoir le (dé)couplage N/P, ou le rôle de Si et Fe.
- le recyclage de la
DOM et l'activité bactérienne - boucle microbienne.
- le transfert
surface - colonne d'eau et le devenir de la matière organique dans l'océan
profond.
Les échelles de temps qui seront abordées
par la station DYFAMED en modélisation sont les suivantes :
- la haute
fréquence (échelle impulsionnelle : apport atmosphérique intense, coup de vent)
le cycle saisonnier
(variabilité des co-limitations, succession d'espèces)
- la variabilité
inter-annuelle (avec la composante anthropique sur le long terme)
Une modélisation déterministe des
co-limitations chimiques successives de la production primaire par la
disponibilité en micro- (Fe) et macro-éléments (sels nutritifs) est proposée
sur la base des expériences d'enrichissement régulières (systèmes LET GO incubés
in situ) et celles prévues au cours des campagnes saisonnières
2003-2004. En outre, une modélisation du processus de diazotrophie et des
effets de la disponibilité de fer et de phosphore sur l'efficacité de ce
processus est envisagée dans la mesure où les travaux menés sur le site DYFAMED
révèlent une fixation significative. La prise en compte de ces co-limitations,
ainsi que l'importance du volet concernant le devenir de la matière organique,
demandent la représentation explicite de plusieurs types de producteurs
primaires. En effet, non seulement certaines espèces sont associées à des
co-limitations spécifiques (e.g., Fe et Si pour les diatomées, Fe et P pour les
diazotrophes, N et/ou P pour le picoplancton), mais la proportion de la
production primaire nette susceptible d'être exportée résulte de la structure
du réseau trophique, fortement conditionnée par le premier échelon. Cette
différenciation permettra en outre de mieux représenter à la fois la boucle
microbienne dans la couche euphotique, régénérant la matière organique
localement, et l'export de flux particulaire, associé à des tailles de
particules différentes. De même, le potentiel d'export de la DOM, très
important dans les régions oligotrophes, nécessite de décomposer le réservoir
générique "DOM" en compartiments caractérisés par leur composition
chimique (rapport C:N:P) et leur degré de labilité (temps de turnover) (Lévy et
al., 1998a). A cet égard, le rôle des TEP dans le bilan de carbone devra
être exploré dans la mesure du possible (Mari et al., 2001). Enfin, pour
décrire le devenir de la matière organique exportée dans les ouches profondes,
le couplage entre un modèle de surface et un modèle de zone aphotique est
prévu. Contrairement aux modèles pélagiques, ces modèles de colonne d'eau sont encore
peu développés et ne prennent en compte que peu de processus. Les premières
études se baseront sur des modèles déjà existants (Kriest & Evans, 1999;
Athias, 2001), et chercheront à les améliorer, en prenant appui à la fois sur
les données de DYFAMED mais aussi sur d'autres programmes d'observation (POMME)
et d'autres bases de données (BATS, HOT).
Les paramétrisations développées seront
incluses dans des modèles biogéochimiques d'écosystème pélagique couplés à des
modèles hydrodynamiques unidimensionnels (modèles OPA ou Symphonie).
Ainsi, un modèle biogéochimique, initialement couplé à la version
tridimensionnelle de Symphonie a déjà permis d'étudier l'impact des
phénomènes physiques de méso-échelle sur l'exportation de carbone dans le Golfe
du Lion (Diaz, 2000). Ce modèle décrit, dans sa configuration actuelle, les
cycles pélagiques de l'azote et du phosphore avec notamment une co-limitation
de la croissance phytoplanctonique par la disponibilité en phosphate et en
nitrate. D'autres études se sont intéressées à l'impact de facteurs physiques à
méso-échelle sur la floraison printanière en Méditerranée nord-occidentale
(Lévy et al., 1998b, 1999, 2000; Mémery et al., 2002; Olivier,
2001).
D'autres paramètres et/ou des
modifications de structure du modèle seront introduits en fonction des
observations de terrains acquises dans le cadre du projet (e.g., variabilité
impulsionnelle des concentrations de substances nutritives, groupes
phytoplanctoniques dominants, etc.). A cet égard, la quantité et la qualité des
données DYFAMED ont déjà permis de mener des exercices de "calibration
objective" de modèles biogéochimiques, grâce àl'écriture du code adjoint
de ces modèles (Faugeras et al., 2002). En effet, les modèles
biogéochimiques sont caractérisés par un nombre important de paramètres
internes, souvent mal connus soit parce que leur mesure est difficile, soit
parce qu'ils représentent plusieurs processus et qu'ils n'ont pas d'équivalents
observables. La calibration de ces modèles est donc délicate. L'assimilation
variationnelle de données est un outil mathématique puissant pour aborder ce
type de problématique. L'acquisition de données à des fréquences régulières au
site DYFAMED fournira les conditions idéales pour ces méthodologies et
quantifier, par exemple, la variabilité saisonnière de certains paramètres, ou
la cohérence des modèles et des observations.
La validation/calibration de ces modèles
se fera donc essentiellement sur sa capacité à reproduire les successions
saisonnières i) des éléments chimiques qui contrôlent le développement de
l'échelon primaire, ii) des groupes fonctionnels phytoplanctoniques contrôlant
l'exportation de carbone au site DYFAMED, et iii) des observations recueillies
dans les pièges à particules (flux de masse, composition).
Sur des échelles de temps de l'ordre de la
saison, le site DYFAMED peut être considéré comme un système 1D vertical. Cette
caractéristique permet d'envisager avec confiance une modélisation 1D lors
d'une expérience de fertilisation. Les zones d'ensemencement en fer dans le
Pacifique équatorial (IRONEX – Coale et al., 1996) et l'Océan Austral
(SOIREE – Boyd et al., 2000) étaient caractérisées par des vitesses de
surface dépassant 50 cm s-1, ce qui est d'un ordre de grandeur plus
élevé que les vitesses moyennes au site DYFAMED. Cependant, les structures de
moyenne et petite échelles sont ubiquistes dans l'océan, et peuvent avoir des
impacts locaux importants, comme cela a été montré par exemple à BATS (McNeil et
al., 1999). En outre, les instabilités du courant Liguro-Provençal peuvent
parfois atteindre une extension suffisante pour toucher le site DYFAMED. Par
conséquent, une modélisation 3D régionale à petite échelle doit être aussi
considérée, ce qui permettra en outre deprendre en compte les échanges
côte-large.
Sur des échelles pluri-annuelles, l'hypothèse 1D n'est plus
valide, ce qui demande alors de vraiment prendre en compte en compte les
apports latéraux. Ceci ne peut se faire que dans des simulations plus globales
de la Méditerranée. Des techniques d'emboîtement sont maintenant
opérationnelles pour effectuer des zooms à haute résolution sur la zone DYFAMED
: les conditions aux limites ouvertes sont alors prescrites par le modèle
grande échelle (Tusseau-Vuillemin et al., 1998; Echevin et al.,
2001). Ce type d'approche est indispensable si on s'intéresse à la variabilité
du signal profond sur des échelles longues, associées par exemple aux
perturbations anthropiques.
Un point très important de ce projet est
que, dans une configuration donnée de limitation, les données acquises
permettront de suivre les évolutions des rapports stoéchiométriques du matériel
exporté. Le couplage production-exportation de C dépend non seulement du
rapport f (production nouvelle/production totale) et de la respiration, mais
aussi de la stoechiométrie C:N:P du matériel exporté par rapport à celle du
phytoplancton (généralement supposée identique, ce qui est manifestement
incorrect). A partir des variations de stoechiométrie sur la verticale et dans
le temps caractérisées au site DYFAMED, sera développée une autre approche,
avec un modèle qui contraint l’exportation verticale de carbone de la zone
euphotique au moyen des valeurs de :
i) production primaire nette,
ii) respiration hétérotrophe,
iii) fixation nette de C,
iv) composition chimique du matériel particulaire et dissous
exporté (C:N ou C:P).
Ce modèle associe donc dans un même
ensemble d'équations deux perspectives jusqu'à présent indépendantes, à savoir
que l'exportation est contrôlée par (1) l'apport d'un élément limitant (dans sa
forme la plus simple, le modèle d'Eppley & Peterson, 1979) et (2) la
différence entre la production primaire nette et la respiration hétérotrophe.
Ce modèle pourrait fournir un cadre conceptuel pour l'exploitation des données
acquises au site DYFAMED.
La
découverte récente que la stoéchiométrie des éléments chimiques (C, N, P, Si)
n’est pas conforme aux rapports constants C:N:P = 106:16:1 de Redfield est une
conséquence inattendue des programmes de recherches pluridisciplinaires tels
que U.S. JGOFS et WOCE (Hood, 2001 ; Michaels et al., 2001). Cette
conclusion provient de l’analyse des mesures réalisées aux stations temporelles
HOT, BATS et DYFAMED, au cours de ces dix dernières années.
Les modèles écologiques basés sur le concept de Redfield
n’arrivent pas à reproduire certains aspects importants du cycle des éléments.
Cela provient essentiellement du fait que les couplages dynamiques entre les
cycles ne sont pas considérés (Karl et al., 2000).
L’approche stoechiométrique a permis la réalisation d’études
expérimentales ou de modélisation (Elser & Hassett, 1994 ; Anderson,
1997 ; Touratier et al., 1999a, b; Touratier et al.,
2000 ; Touratier et al., 2001). Avec ces études, il est possible
d’entrevoir l’importance et surtout les conséquences de la variabilités de
rapports C:N:P:Si au niveau des composants et des processus sur la dynamique
des écosystèmes marins. Le principal avantage de cette approche réside dans sa
capacité à intégrer et simuler le cycle biogéochimique des principaux éléments
(Sterner & Hessen, 1994 ; Elser et al., 1996). Par cette
méthode, ce n’est pas seulement la quantité, mais aussi la qualité (les
rapports des éléments) des composés qui influencent un nombre important de
processus au sein des réseaux trophiques. Ces aspects nouveaux ont stimulés
l’émergence d’une nouvelle génération de modèles d’écosystème (Anderson,
1997 ; Touratier et al., 2000, 2001) où la variabilité des rapports
élémentaires est explicitement parametrisée. Le but de cette étude, commune aux
propositions PROOF MELISSA et ACTION (Anthropogenic Carbon : Temporal
Increase, Observations and Numerization; proposé par C. Goyet), est de fournir
un modèle stoéchiométrique capable de décrire la dynamique des principaux
compartiments inorganiques, organiques, particulaires et dissout au site
DYFAMED.
La Figure 5 montre la structure du modèle qui sera construit
pour simuler la dynamique du réseau trophique au site DYFAMED. Pour construire
un tel modèle avec les unités C, N, P et Si, les paramétrisations de Anderson
(1997) et Touratier et al. (2001) décrivant les processus autotrophiques
et hétérotrophiques, seront intégrées au modèle d’écosystème construit par
Touratier et al. (en prép.).
Dans le cadre du projet ACTION, l'objectif sera de calibrer et
de valider la structure du modèle stoéchiométrique éléments qui sera couplé à
un modèle de chimie du carbone. Ce
modèle est destiné a être couplé au modèle OPA 3D du LODYC pour toute la
Méditerranée. Dans le cadre de MELISSA, le but sera d’étudier l’influence des
concentrations et des rapports des nutriments (nitrates/nitrites, ammonium,
phosphates, silicates) sur la composition et la dynamique des réseaux trophiques et les effets
rétro-actifs sur le type d’élément qui limite la croissance du phytoplancton.
Nous analyserons aussi les changements de limitation (N vers P ou vice-versa)
observés au site DYFAMED (Marty et al., 2002; voir Figure 3).
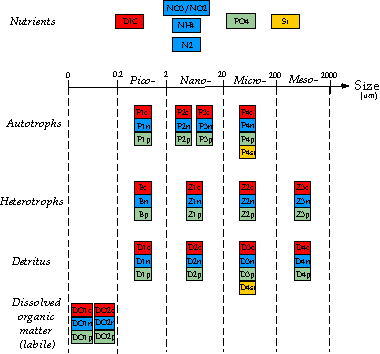
|
Figure
5: Structure of the stoichiometric model with the highest level of
complexity, to be build up for the MELISSA and ACTION projects. Red is for C,
blue is for N, green is for P, and yellow is for Si. For clarity, processes
between variables are not represented. Note that two types of nano-autotrophs
are considered (non-calcifying and calcifying species; i.e. P2 and P3,
respectively). Two types of dissolved organic matter are also considered,
i.e. the low and high molecular
weight compounds (DO1 and DO2, respectively). |
Collaborations
Ru Cheng Tian (Dept. of Earth and Planetary
Sciences, Harvard University, USA)
IV.2.
Evolution du système/Modèles prédictifs
La
variabilité des rapports P:Si:N:C à l’échelle saisonnière sur le site DYFAMED
sera étudiée en intégrant les séries temporelles déjà disponibles. Pour le
matériel particulaire, le suivi du rapport C:N:Si:P dans la matière vivante
(matériel filtré) et dans les pièges à particules (200 m) sera pris en compte.
Pour le matériel dissous, les concentrations de nutriments (NO3-,
Si(OH)4-, PO43-) seront prises en compte.
La modélisation permettra d'évaluer l’impact des apports
externes sur les rapports molaires et les flux de matière dans la colonne
d’eau. L’étude de la distribution des espèces zooplanctoniques pourrait apporter
des éléments de de réponse sur les conséquences trophiques d’un éventuel
déséquilibre des rapports N:Si:P. Cependant, il n’y a pas de valeurs de
référence aujourd’hui qui permette d’envisager une évolution. De telles
observations au site DYFAMED pourraient toutefois servir de référence pour les
évolutions futures.
D'une part, de nombreux processus fondamentaux interagissent,
très souvent d’une manière non linéaire, avec des échelles de variabilité qui
recouvrent plusieurs ordres de grandeur, ce qui pose problème au niveau de
l’observation. D'autre part, l’activité humaine peut non seulement fortement
perturber flux et bilans de constituants biogéochimiques, mais aussi modifier
le comportement d’écosystèmes dans leur globalité. Les relations entre ces paramètres
étant souvent de nature non linéaires, voire chaotiques, des modèles de type
“ réseaux de neurones ” seront utilisés dans le projet. Ils
présentent l’avantage d’intégrer les processus stochastiques du comportement de
l’écosystème, mais en contrepartie nécessitent de nombreuses données. Les
séries temporelles du site atmosphérique du Cap Ferrat et du site marin
DYFAMED, et l'acquisition des données dans le cadre du projet permettront une
utilisation optimale de ce type de modélisation prédictive. Les résultats
obtenus avec de tels modèles pourront être comparés avec ceux des modèles à
compartiments plus classiques. Le projet MELISSA pourrait également être aussi
l’occasion d’utiliser des méthodes d’analyses multivariées non linéaires pour
examiner les relations entre les différentes variables prises en compte.




