...... click to go to ......
II.1.1 Les dťterminations pigmentaires
II.1.2 Diversitť et abondance du
picoplancton procaryote et eucaryote
II.1.3
Mesures de l'expression de gŤnes de cyanobactťries en milieu ocťanique
II.1.5
Exercice d'intercalibration des compteurs Hiac et Coulter.
II.1.6
Mesure des flux d'azote.
II.1.7
Amťlioration de la mťthode de dťtermination des orthophosphates.
II.2 Annťe 2004 : Campagne
BIOSOPE dans le Pacifique Sud
II.2.1
Trajet thťorique de la campagne
II.2.2
Mesures effectuťes ŗ bord
II.2.3
Possibilitť d'avoir un deuxiŤme bateau (Golden Shadow) sur site
II.3 Annťe 2004-2005 (et
suivantes): Traitement de la campagne & synthŤse sur l'oligotrophie
II.1
Annťes 2002, 2003 & 2004: Dťveloppement
mťthodologiques en amont, contrŰle de qualitť des donnťes
La programmation ťventuelle d'une campagne
ocťanographique dans la zone a priori la plus oligotrophe de la
planŤte pose un certain nombre de questions et de problŤmes quant
aux mťthodes qui seront mises en œuvre. En effet, oligotrophie
signifie souvent "traces" au niveau d'un certain nombre
de grandeurs biogťochimiques. Il s'en suit que certaines mťthodes
risquent d'Ítre utilisťes au niveau de leur limite de leur dťtection
actuelle. Pour ťviter de "mauvaises surprises" (e.g. incapacitť
ŗ mesurer un certain nombre de paramŤtres), ce problŤme doit Ítre
identifiť a priori, et les moyens donnťs afin d'Ítre opťrationnels
lors de la campagne. Un travail mťthodologique visant ŗ dťvelopper
des mťthodes plus sensibles ou ŗ accroÓtre la sensibilitť des mťthodes
actuelles devra donc Ítre entrepris. Dans ce cadre, nous retenons
plusieurs axes mťthodologiques qui devront faire l'objet de recherches
soutenue en prťalable ŗ la campagne.
II.1.1 Les dťterminations pigmentaires.
Lors de la campagne PROSOPE, un exercice
d'intercomparaison des mťthodes HPLC d'analyse des pigments avait
ťtť organisť. Cet exercice, impliquait quatre laboratoires (LOV
Villefranche, NASA, JRC Italie, Afrique du Sud). Il fut mis en ťvidence
que l'accord entre les rťsultats des diffťrents laboratoires diminue
lorsque l'on compare des eaux de plus en plus oligotrophes (
Hooker et al. 2001
, Figure 5). Cet exercice d'intercomparaison
fut le premier ŗ utiliser des ťchantillons naturels comme "matťriel
test", des standards de pigments ťtant jusqu'alors les seules
rťfťrences disponibles. Rťcemment (congrŤs NASA-SIMBIOS ŗ Washington
en Janvier 2002), il fut reconnu que ce type d'exercice sur ťchantillon
naturel doit Ítre privilťgiť car c'est le seul permettant d'apprťcier
toutes les sources de variabilitť entre les diffťrents laboratoires.
Nous comptons initier un exercice international de plus grande ampleur
lors de la campagne BIOSOPE : 6 laboratoires seront impliquťs et
24 ťchantillons tests en triplicata seront distribuťs ŗ ces laboratoires
(12 ťchantillons tests en triplicatas avaient ťtť distribuťs ŗ quatre
laboratoire durant PROSOPE). L'organisation de cet exercice d'intercomparaison
de grande ampleur bťnťficiera du recrutement rťcent de Josťphine
Ras comme IE spťcialiste des pigments au LOV de Villefranche. Cette
personne avait participť ŗ l'organisation du premier exercice sur
CDD financť par la NASA. Les financements demandťs serviront ŗ prťparer
les systŤmes de filtration simultanťs et de stockage des ťchantillons.
II.1.2 Diversitť et abondance du picoplancton procaryote
et eucaryote
La diversitť de Prochlorococcus sera ťtudiťe en utilisant une mťthode de PCR-RFLP basťe sur l'utilisation du gŤne pcb (voir plus haut) mťthode qui a dťjŗ ťtť testťe au laboratoire ( Garczarek et al. 2000 ) et lors de PROSOPE (Garczarek, Dufresne et Partensky, en prťp.).
Les mťthodes qui seront utilisťes pour mesurer la diversitť et l'abondance du picoplancton eucaryote sont fondťes sur l'analyse du gŤne de l'ARN ribosomal 18S, marqueur taxonomique universellement utilisť et pour lequel la base de donnťe existante est la plus complŤte ŗ l'heure actuelle. Parmi ces mťthodes, l'une, le FISH (utilisation de sondes fluorescentes dťtectťes par microscopie), est maintenant parfaitement au point et utilisable en routine (Not et al. en rťvision). De plus un grand nombre de sondes sont dťjŗ disponibles (Tableau 1) Par contre, deux autres approches la DGGE et la PCR quantitative sont encore en cours de dťveloppement.
Tableau 1: ARNr 18S probes available for in situ hybridization.
| Target |
Autotroph/ |
Probe name |
| Euk |
|
Euk1209R |
| Chlorophyta |
A |
CHLO02 |
| Prasinophyceae Pseudocourfeldiales |
A |
PRAS01 |
| Prasinophyceae Prasinococclaes |
A |
PRAS03 |
| Prasinophyceae Mamiellales |
A |
PRAS04 |
| Praisinophyceae Clade CCMP1205 |
A |
PRAS05 |
| Praisinophyceae Clade OLI11305 |
A |
PRAS06 |
| Stramenopiles (Heterokonta) |
A/H |
HETER01 |
| Bolidophyceae |
A |
BOLI01 |
| Dictyochophyceae |
A |
DICTYO01 |
| Eustigmatophyceae |
A |
EUSTI01 |
| Pelagophyceae |
A |
PELA01 |
| Stramenopiles novel clade III |
H |
NS03 |
| Stramenopiles novel clade IV |
H |
NS04 |
| Stramenopiles novel clade VII |
H |
NS07 |
| Dinophyceae + Apicomplexa |
A |
DINO_B |
| Haptophyta |
A |
PRYM02 |
| Cryptophyceae |
A |
CRYPT12 or CRYPT07 |
∑ DGGE (ARNr 18S)
La DGGE permet de sťparer par ťlectrophorŤse des fragments dont la taille est identique mais dont la sťquence est diffťrent. Le principe en est le suivant ( Figure 6):
3)
En fonction de leur sťquences, les fragments amplifiťs vont migrer
plus ou moins loin sur le gel et on obtiendra in
fine une image de la diversitť du peuplement initial (Figure
6).
4)
Les fragments peuvent ensuite Ítre clonťs et sťquencťs
afin de connaÓtre l'identitť taxonomique des diffťrentes bandes.
|
Figure 6: Principe de la DGGE. Analyse de l'ťvolution sur 57 jours des populations bactťriennes associťs ŗ Alexandrium tamarense. Chaque bande correspond ŗ un taxon diffťrent et les bandes indiquťes en rouge ont ťtť clonťes et sťquencťes afin de dťterminer leur identitť. Par exemple la bande 43 correspond ŗ une protťobactťrie beta (mťthylotrophe) alors que la bande 44 est une Cytophagale (Vaulot et al, en prťpa). |
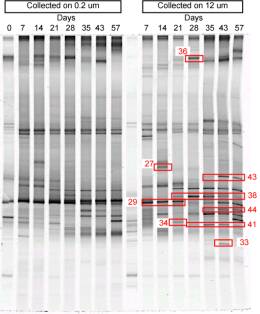
Le groupe de Roscoff possŤde dťjŗ une certaine expťrience de cette technique car elle a ťtť appliquťe rťcemment ŗ l'analyse des populations bactťriennes associťes au dinoflagellť Alexandrium tamarense (Vaulot et al. en prťp.). Elle est actuellement utilisťe pour les eucaryotes par des collŤgues espagnols ( Diez et al. 2001 ) avec lesquels une collaboration est en cours pour implanter cette technique ŗ Roscoff.
∑
PCR quantitative (ARNr 18S)
II.1.3 Mesures de l'expression de
gŤnes de cyanobactťries en milieu ocťanique
Les mťthodes de mesures classiques de l'expression des gŤnes (en particulier Northern blot) sont difficiles ŗ utiliser sur des ťchantillons naturels car elles rťclament beaucoup de matťriel. Une mťthode particuliŤrement attractive est la RT-PCR quantitative. La mťthode consiste ŗ transcrire les ARN messagers en ADN puis ŗ amplifier l'ADN avec des amorces spťcifiques du gŤne ciblť. La rťaction de PCR est suivi en temps rťel par fluorescence (voir ci-dessus). Cette mťthode est rapide, quantitative et peut Ítre appliquťe sur un grand nombre d'ťchantillons simultanťment. Elle est utilisťe par le groupe de Roscoff sur des cultures et vient d'Ítre appliquťe par des collŤgues allemands ŗ des ťchantillons naturels de Prochlorococcus (Holtzendorff et al. soumis). Ces techniques sur ťchantillons naturels seront donc dťveloppťes au cours de deux prochaines annťes par le groupe de Roscoff. Une ťtape importante est la mise au point d'amorces suffisamment gťnťrales pour cibler l'ensemble des populations naturelles de Prochlorococcus. Afin de mettre au point ces amorces, il faut d'abord sťquencer le gŤne ciblť (par exemple groEL pour les gŤnes de rťponse au stress) dans le plus grand nombre possible d'ťchantillons et utiliser cette base de donnťe de sťquences pour trouver les zones du gŤne les plus conservťes. Ces travaux prťliminaires seront effectuťs au cours de deux premiŤres annťes du projet BIOSOPE.
II.1.4 Les mťthodes optiques
ō
La matiŤre organique dissoute colorťe
(CDOM en anglais) est une quantitť difficile ŗ mesurer dans les
eaux ocťaniques (cas I) : sa concentration est gťnťralement proche
des limites de dťtection des mťthodes optiques classiques. Certes,
le CDOM absorbe plus dans l'UV que dans le visible, et cette caractťristique
peut parfois Ítre utilisťe pour mesurer "facilement" le
CDOM. Toutefois il reste absolument nťcessaire de rťaliser des mesures
quantitatives et prťcises de CDOM dans le visible car seules ces
mesures sont "exploitables" pour rťaliser un bilan ("closure"
en anglais) de la contribution de toutes les substances aux propriťtťs
optiques des eaux; cette ťtape est nťcessaire aussi bien pour interprťter
la couleur de l'eau que pour quantifier (ou modťliser) l'ťclairement
disponible pour la photosynthŤse. Pour amťliorer la dťtection du
CDOM, il faut augmenter la longueur du trajet optique : les limites
de dťtection sont alors rťduites proportionnellement ŗ cette augmentation.
Les mťthodes spectrophotomťtriques classiques permettent l'emploi
de cuves de 10 cm et sont gťnťralement insuffisantes. Le AC9 (tubes
de 25 cm) est une rťelle amťlioration, mais ces mesures continues
nťcessitent d'Ítre validťes par des mťthodes discrŤtes. Tout rťcemment
( D'sa et al. 1999 ; D'sa and Steward 2001 ) ont proposť d'utiliser le principe
des cellules de mesures capillaires ŗ paroi rťflťchissante (Liquid
Waveguide Capillary cell) de longueur variable (jusqu'ŗ 2 m) pour
mesurer le CDOM. Ces recherches ont abouti au dťveloppement d'un
appareil prototype, l'Ultrapath (commercialisť par World Precision
Instrument). Nous sommes en relation avec la compagnie qui pourrait
nous proposer pour emprunter un prototype pour rťaliser des tests
en vue d'une utilisation ťventuelle lors de la campagne BIOSOPE.
ō
En 1970, Elterman introduisait
la ę cavitť intťgrante Ľ (ou ę integrating-cavity
absortion meter Ľ : ICAM) pour la mesure de faibles absorptions
par des fluides. Cette cavitť est consiste en une sphŤre aux parois
blanches fortement diffusantes, dans laquelle la lumiŤre est amenťe
de maniŤre homogŤne. Pour la mesure d’absorption, cette cavitť
est remplie par l’ťchantillon. Cette technique fut reprise
et amťliorťe par Pope and Fry (1997 ) afin de mesurer avec grande prťcision
le spectre du coefficient d’absorption de l’eau pure.
Cette prťcision est rendue possible par les trŤs longs chemins optiques
qui rťsultent des rťflexions nombreuses sur les parois (alors que
la cavitť fait la taille d’une boule de pťtanque !). Les mesures
de Pope and Fry (1997 ) font aujourd’hui rťfťrence.
Kirk (1997 ) a proposť une version modifiťe de
la cavitť intťgrante (ę point-source integrating-cavity meter Ľ :
PSICAM) qui permet de mesurer ŗ la fois de faibles et de fortes
absorptions par l’eau de mer naturelle. Cette technique est
idťale pour les ťchantillons naturels parce que la cavitť intťgrante
a cet avantage supplťmentaire que sa mesure de l’absorption
n’est en aucune faÁon altťrťe par la diffusion particulaire
(
Leathers et al. 2000
). En effet, la prťsence de particules
ne peut rendre le champ radiatif plus diffus qu’il ne l’est
dťjŗ dans la sphŤre du fait des parois diffusantes (en spectralon).
Cette technique est donc idťale pour la campagne BIOSOPE parce qu’elle
offre (1) la sensibilitť nťcessaire pour ces eaux ultra-oligotrophes
et (2) elle permet une mesure rigoureuse de l’absorption par
les substances dissoutes et particulaires ( Pope et al. 2000 ). Parce qu’il n’existe aucune version commerciale de cet instrument,
un PSICAM sera dťveloppť par l’ťquipe d’optique et de
tťlťdťtection du LOV.
ō
Finalement, pour la mesure de la fluorescence
variable sur les ťchantillons discrets, il sera nťcessaire d’apporter
des modifications au PAM afin d’en amťliorer la sensibilitť.
Durant la campagne PROSOPE, il fut difficile d’obtenir des
mesures fiables prŤs de la surface, lŗ oý la concentration en chlorophylle
est la plus faible du fait de la forte carence en nutriments et
de la photoacclimatation (
Bruyant 2002
). Or, cette partie de la colonne d’eau
prťsente un intťrÍt particulier parce qu’on y rencontre les
stress environnementaux les plus accentuťs. La modification du PAM
comprendra (1) le remplacement du dťtecteur (photo-diode remplacťe
par un photomultiplicateur sensible dans le rouge et refroidi par
effet Pelletier), (2) la modification du porte-cuvette afin d’amťliorer
la rťcolte des photon ťmis par fluorescence et, ťventuellement,
(3) le remplacement de la source d’excitation (lampe xťnon
remplacťe par une grappe de LED). Ces modifications seront apportťes
par le fabricant du PAM (Walz) selon les recommandations de l’ťquipe
d’optique et de tťlťdťtection du LOV.
II.1.5 Exercice d'intercalibration des compteurs Hiac et Coulter.
Le compteur optique Hiac (Pacific Scientific) s'est avťrť Ítre un outil fiable et robuste pour des opťrations soutenues ŗ la mer (Almofront Claustre et al. 2000 , Prosope Oubelkheir 2001 et Pomme), capable de produire un volume d'information considťrable, correspondant aux besoins d'investigations prolongťes et ŗ haute frťquence induits par la comprťhension des processus biogťochimiques affectant l'ensemble des particules en suspension. Nťanmoins, un examen approfondi des distributions de taille obtenues avec cet appareil laisse pressentir une dťformation de la forme des spectres (dont l'origine reste ŗ ťlucider: conformation gťomťtrique de l'optique, indice de rťfraction des particules marines, parts respectives des signaux absorption et de diffusion impliquťes dans le signal d'attťnuation, etc.) pour des tailles voisines de 2,5 Ķm.
L'utilisation conjointe d'un autre compteur, comme le Coulter (Coultronics), fonctionnant sur un principe diffťrent, est complťmentaire de celle du Hiac. En effet, contrairement ŗ ce dernier, le mode opťratoire du Coulter ne lui permet pas d'Ítre utilisť in situ pour l'acquisition d'un volume d'information aussi important. Par contre, les spectres produits par le Coulter semblent corrects et permettraient, sinon de comprendre le biais produit par le Hiac, du moins de le corriger; l'association des deux compteurs, procurant des donnťes ŗ la fois nombreuses et non biaisťes, est donc complťmentaire.
Un exercice prťalable d'intercomparaison des deux appareils (dans l'hypothŤse ou le financement d'un coulter counter pour le LOV de Villefranche serait soutenu en 2002 par la CSOA) est donc nťcessaire, qui requiert l'utilisation de sphŤres de tailles et d'indice de rťfraction diffťrents et connus.
II.1.6 Mesure des flux d'azote.
Les mesures directes de la fixation d’azote (ŗ l’aide de l’isotope stable 15N) sont encore peu nombreuses ŗ l’heure actuelle, la plupart des estimations ayant ťtť jusqu’ŗ prťsent obtenues par une mťthode indirecte (dite ŗ l’acťtylŤne) qui nťcessite l’utilisation de facteurs de conversion. La technique proposťe ici est basťe sur celle dťveloppťe par Montoya et al. (1996 ) pour laquelle l’addition de traceur se fait sous forme gazeuse et perturbe au minimum l’ťchantillon. Ces mesures se feront sur la fraction totale en utilisant des filtres GF/F et 0.2 Ķm Anopore. Ces derniers filtrent montrent une meilleure capacitť de rťtention de la matiŤre dans les eaux oligotrophes ( Raimbault et al. 1999 ), mais sans que cela entraÓne une diffťrence significative sur les taux de prise de nitrate. Dans la mesure oý se rťsultat pourrait Ítre diffťrent pour la fixation d’azote il est nťcessaire de le vťrifier.
Une ťtude particuliŤre portera sur la fraction nanoplanctonique (>3 Ķm) par l’utilisation de filtre de porositť 3Ķm en argent compatible avec une analyse au spectromŤtre de masse. Ceci permettra d’isoler la fraction nanoplanctonique et de rťduire la dilution isotopique en ťliminant les particules de petites tailles qui ne seraient pas impliquťes dans le processus de fixation d’azote (donc peu ou pas enrichies en azote-15). C’est une maniŤre d’amplifier le signal de ce processus dont les taux sont certainement trŤs faibles et concernent un trŤs faible nombre de cellules par litre ( Zehr et al. 2001 ). Les premiers tests de cette mťthode auront lieu en 2002 sur les campagnes actuellement programmťes (e.g. DIAPALIS).
II.1.7
Amťlioration de la mťthode de dťtermination des orthophosphates.
Les mťthodes pour la dťtermination des orthophosphates qui ont commencť ŗ Ítre employťes dans le cadre de PROSOPE, notamment la mťthode MAGIC et la mťthode indirecte utilisant l’isotope RA du phosphore (33P), nťcessitent d’Ítre amťliorťes. Les dťveloppements rťcents de la mťthode MAGIC suggŤrent de remplacer la centrifugation par une simple filtration du prťcipitť dont l’efficacitť dans l’adsorption des orthophosphates semble quantitative pour des concentrations de soude ajoutťe nettement infťrieures ŗ celles prťconisťes auparavant ( Thomson-Bulldis and Karl 1998 , Wu et al. 2000 ). Cette amťlioration devrait permettre, d’une part de baisser la limite de dťtection des orthophosphates et d’autre part, d’envisager une utilisation plus systťmatique de cette mťthode. Il est ťgalement prťvu de prendre en compte l’interfťrence de l’Arsenate gťnťralement nťgligťe jusqu’alors.
II.2
Annťe 2004 : Campagne BIOSOPE dans le Pacifique
Sud
Les objectifs scientifiques de cette campagne ont ťtť exposťs plus haut. Le prťsent projet n'est pas le projet de campagne (celui-ci sera soumis en temps voulu ŗ l'IFREMER, c'est ŗ dire en Janvier 2003). Il est actuellement relativement inťgal quant aux diffťrentes orientations de cette campagne et doit Ítre considťrť comme prťliminaire; il sera affinť lors de rťunions entre les diffťrents participants durant l'annťe 2002. On peut nťanmoins donner un certain nombre d'indications quant au trajet prťvu et ŗ l'organisation des mesures ŗ bord.
II.2.1 Trajet thťorique de la campagne
La campagne BIOSOPE devrait idťalement avoir lieu entre novembre et fťvrier:
∑ Le maximum de biomasse dans le panache des Óles marquises est observť entre aoŻt et dťcembre ( Signorini et al. 1999 ) mais en Janvier et Fťvrier l'imagerie SeaWifs montre ťgalement des biomasses significatives.
∑ L'oligotrophie du tourbillon est maximale entre novembre et mars (climatologie SeaWiFS)
†††††††††††
La figure 7 illustre le trajet thťorique de la campagne BIOSOPE correspondant ŗ une demande de temps bateau d'environ 54 jours (calculť pour une vitesse nominale ŗ 11 nœuds). Le tableau 2 rťsume les ťtapes principales de cette campagne.
Les quatre grands types d'ťtudes menťes durant cette campagne seront les suivantes :
∑ Filament upwelling : Il s'agira vraisemblablement d'une ťtude de 7 stations rťparties le long d'un filament d'upwelling (extension de 300-400 km ŗ partir du cœur). Chaque station durerait une vingtaine d'heures. Le choix de la zone sera arrÍtť en dťbut de campagne ŗ partir d'imagerie couleur de l'eau (SeaWiFS et MERIS).
∑
Tourbillon 1 et 2 : Il s'agira de deux ťtudes en point
fixe de 5 jours. La station "tourbillon 1" devrait idťalement
Ítre placťe sur la bordure du tourbillon du Pacifique Sud. La station
tourbillon 2 serait au centre, lŗ oý l'oligotrophie "extrÍme"
est attendue.
∑ Panache Marquises : MÍme principe que pour l'ťtude du filament d'upwelling: 7 stations d'une vingtaine d'heures rťparties selon une direction Ouest Nord-Ouest. Positionnement du rťseau gr‚ce ŗ l'imagerie couleur.
∑ Les 21 stations "courtes" de 6 heures permettent de caractťriser les gradients entre ces extrÍmes: elles seront occupťes autour du midi solaire.
Tableau 2: Theoretical scenario of BIOSOPE cruise.
Transit times are calculated for a ship speed of 11 knots. Long
stations are identified by bold characters, while departure and
arrival ports are underlined. For latitude and longitude, decimals
correspond to tenth of degrees.
|
|
Latitude Sud |
Longitude †Ouest |
Distance (miles) |
Temps Transit (jours) |
Temps en Station (jours) |
|
|
|
|
|
|
|
| Valparaiso
(Chili) |
33.0 |
71.5 |
|
|
1 |
| Filament Upwelling |
27.0 |
74.0 |
386 |
1.46 |
7.0 |
| st1 |
27.0 |
77.5 |
198 |
0.75 |
0.3 |
| st2 |
27.0 |
81.0 |
198 |
0.75 |
0.3 |
| st3 |
27.0 |
84.5 |
198 |
0.75 |
0.3 |
| Tourbillon 1 |
27.0 |
88.0 |
198 |
0.75 |
5.0 |
| st4 |
27.0 |
91.5 |
198 |
0.75 |
0.3 |
| st5 |
27.0 |
95.0 |
198 |
0.75 |
0.3 |
| st6 |
27.0 |
98.5 |
198 |
0.75 |
0.3 |
| st7 |
27.0 |
102.0 |
198 |
0.75 |
0.3 |
| st8 |
27.0 |
105.5 |
198 |
0.75 |
0.3 |
| st9 |
27.0 |
109.0 |
198 |
0.75 |
0.3 |
| Ile de P‚ques |
27.0 |
110.0 |
57 |
0.21 |
2.0 |
| st10 |
27.0 |
112.0 |
113 |
0.43 |
0.3 |
| Tourbillon 2 |
27.0 |
115.5 |
198 |
0.75 |
5.0 |
| st11 |
27.0 |
119.0 |
198 |
0.75 |
0.3 |
| st12 |
27.0 |
122.5 |
198 |
0.75 |
0.3 |
| st13 |
27.0 |
126.0 |
198 |
0.75 |
0.3 |
| st14 |
27.0 |
129.5 |
198 |
0.75 |
0.3 |
| st15 |
27.0 |
133.0 |
198 |
0.75 |
0.3 |
| st16 |
27.0 |
136.5 |
198 |
0.75 |
0.3 |
| st17 |
27.0 |
140.0 |
198 |
0.75 |
0.3 |
| st18 |
23.7 |
140.0 |
198 |
0.75 |
0.3 |
| st19 |
20.4 |
140.0 |
198 |
0.75 |
0.3 |
| st20 |
17.1 |
140.0 |
198 |
0.75 |
0.3 |
| st21 |
13.8 |
140.0 |
198 |
0.75 |
0.3 |
| Panache Marquise |
10.0 |
140.0 |
228 |
0.86 |
7.0 |
| Tahiti |
18.0 |
150.0 |
750 |
2.84 |
1 |
|
|
|
|
|
|
|
| Total |
|
|
5894 |
22 |
32 |
II.2.2 Mesures effectuťes ŗ bord [1]
∑ Mesures continues :
ō A partir de la rosette : CTD, fluorescence, Fluorescence variable,
ō A partir de l'eau pompťe : CTD, Fluorescence in vivo, pCO2, spectre de taille
ō A partir d'air pompť (mesures semi continu) : composition de l'aťrosol (voir remarque)
ō Profileurs optiques: propriťtťs inhťrentes (absorption dissoute et particulaire, attťnuation et retro diffusion) et apparentes (ťclairement spectral ascendant et descendant, radiances ascendantes)
∑ Mesures discrŤtes (eau de la rosette)
ō Mesures de stock : Nutritifs, Fe, Silice lithogťnique et biogťnique, MOD, MOP (Figure 8), TC02, pigments, mycosporine amino-acids, cytomťtrie (picoplancton photosynthťtique, bactťries, virus), abondance et spectre de taille des particules, taxonomie phytoplanctonique
ō Mesures de flux : production primaire (Figure 9), assimilation d'azote et de phosphore, fixation de N2, production de silice biogťnique, cinťtiques d'absorption de l'acide silicique, courbes P vs E.
ō Mesures de la diversitť gťnťtique des cyanobactťries (Prochlorococcus) et de la biodiversitť taxonomique des eucaryotes (DGGE, PCR quantitative, FISH)
ō Analyse de l'expression de gŤnes liťs ŗ au stress et la photoprotection chez Prochlorococcus
ō Mesures optiques : absorption particulaire et dissoute, fluorescence variable (PAM).
∑ Trappes ŗ sťdiment dťrivantes (stations longues) : paramŤtres de base (C, N, P), marqueurs organiques et ťlťments traces (ce point reste encore ŗ ťclaircir)
Remarque : Actuellement la composition de l'ťquipe a une forte composante "ocťan" et assez faible du point de vue de l'atmosphŤre. Des contacts seront pris au cours de l'annťe 2002, pour "intťresser" des spťcialistes de la basse atmosphŤre pour des mesures permettant de mieux caractťriser l'aťrosol au dessus des zones explorťes.
II.2.3 Possibilitť d'avoir un deuxiŤme bateau (Golden Shadow) sur site
Le Golden Shadow, bateau de 67 m appartenant ŗ un mťcŤne ťtranger, a pour principale vocation la rťalisation de films scientifiques. Il est nťanmoins "disponible" ŗ la communautť scientifique moyennant l'acceptation de projets par un comitť scientifique international prťsidť par le Professeur J. Jaubert. Ce bateau peut Ítre trŤs utile, notamment pour des expťriences d'optique in situ lťgŤre nťcessitant la mise ŗ l'eau manuelle de profileurs ŗ partir du bateau ou mieux d'annexes (le Golden Shadow est trŤs pratique pour la mise ŗ l'eau de telles embarcations). Nous envisageons ťventuellement (si le Golden Shadow a des "chances" de se trouver dans le Pacifique durant la pťriode pressentie pour la campagne Atalante) de faire une demande de temps bateau, au moins pour un LEG. Une partie des 14 places disponibles pourrait Ítre alors mis ŗ la disposition de cinťastes, l'autre partie permettant d'accueillir des mesures d'optique in situ. Des contacts avec le Pr. Jaubert seront pris au cours de l'annťe 2002.
II.3 Annťe 2004-2005 (et suivantes): Traitement de la campagne & synthŤse sur l'oligotrophie
Bien ťvidemment, les annťes 2004, 2005 et les suivantes seront consacrťes ŗ l'exploitation de la campagne.
Plus gťnťralement, ŗ l'issue de la campagne BIOSOPE, la communautť franÁaise disposera d'une connaissance (et d'une base de donnťes) unique sur les systŤmes oligotrophes. En effet, les ťtudes biogťochimiques des grandes zones oligotrophes peuvent Ítre considťrťes comme une "spťcialitť franÁaise". Les campagnes Chlomax (Mer des Sargasses), EUMELI 3 et 4 (Tourbillon subtropical Atlantique Nord), MINOS et PROSOPE (Mťditerranťe orientale) OLIPAC et BIOSOPE (Tourbillon Pacifique Subtropical Sud) ont (auront) toutes abordť et dťcrit les propriťtťs biologiques, biogťochimiques et optiques d'une variťtť de systŤmes oligotrophes. Cette connaissance pourrait Ítre formalisťe dans le cadre d'une synthŤse sur les systŤmes oligotrophes qui, rappelons-le, reprťsentent 40 % de la surface de l'ocťan mondial. Parmi les ťtudes synthťtiques qui devront Ítre entreprises, on peut, pour l'instant, les ťnoncer sous forme de questions (une rťunion en 2002 devrait prťciser le cadre de ces futures synthŤses):
∑ Quels sont les similaritťs et les diffťrences entres les diffťrentes zones oligotrophes et pourquoi?
∑ En particulier, qu'elle est l'importance, pour la manifestation de leurs caractťristiques biogťochimiques, de la proximitť de ces zones vis-ŗ-vis des sources d'aťrosols dťsertiques?
∑ Quelle sont les relations entre la stratification, la pťnťtration de l'ťclairement sous-marin, la position des nutriclines et du maximum profond de chlorophylle? Ces interactions sont-elles modťlisables?
∑ Existe t'il des relations gťnťrales entre propriťtťs photo physiologiques (e.g. Ek) et des grandeurs physiques modťlisables (e.g ťclairement moyen dans la couche superficielle)?
∑ La diversitť microbienne varie-t'elle en fonction du degrť d'oligotrophie?
∑ Y a t'il une relation entre la nature des limitations [Nitrates (dťpendant du Fer) ou Phosphate] et la structure des communautťs autotrophes?
∑
On sait que la relation entre la couleur de l'eau et [Chla]
prťsente des nuances parfois importantes selon les rťgimes oligotrophes
: quelle en est la cause?
Au prťalable, il sera vraisemblablement nťcessaire qu'un effort soit menť sur la mise en forme des donnťes archivťes afin de les mettre dans un format utilisable ŗ une synthŤse sur l'oligotrophie.
Un colloque sur l'oligotrophie pourrait Ítre organisť durant cette phase de synthŤse en invitant, notamment, les acteurs impliquťs dans l'ťtude des systŤmes autour des zones HOTS, BATS et DYFAMED qui apporterait un ťclairage quant ŗ la variabilitť dťcennale en rťgime oligotrophe, variabilitť que le travail de synthŤse menť dans le cadre de BIOSOPE n'aborderait pas.
[1]
En plus des mesures ťnoncťes dans cette
section, deux types d'expťriences pourraient Ítre entreprises.
:
ō
d'une part, des expťriences liťes aux effets des ultraviolets
sur les dommages cellulaires et les modifications de structure
molťculaire dans le compartiment bactťrien [collaboration avec
le projet FEXBIO (Richard Sempťrť)]
ō d'autre part des mesures liťes ŗ l'activitť bactťrienne profonde et utilisant les bouteilles hyperbares dťveloppťes par Armand Bianchi..





