La reconstruction des
variations de la productivité océanique au cours du temps
est étudiée dans le but de comprendre les changements climatiques,
leur influence sur la pompe biologique de carbone, et réciproquement.
Dans ce contexte, des indicateurs (couramment appellés “ proxies ”)
sont développés, qui permettent de quantifier empiriquement
les liens entre la productivité océanique actuelle et la
fabrication, la préservation et le taux de dépôt de
ces indicateurs dans les sédiments. Le but ultime étant de
dégager des relations empiriques dans l'océan actuel pour
les extrapoler ensuite à l'analyse des carottes sédimentaires.
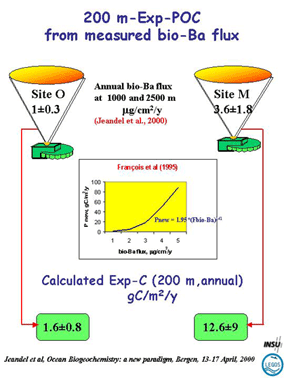 Figure
1 : Flux of export POC, based on biogenic barium fluxes measured
at EUMELI (values on top, on the left for the oligotrophe site
and the right for the mesotrophe site). We applied François
et al. (1995) algorithm to these fluxes and obtained POC fluxes
(values in green squares) in agreement with results acquired with
other approaches (total primary production, drifting traps, POC
fluxes in steady traps etc…). From Jeandel et al., 2000.
Figure
1 : Flux of export POC, based on biogenic barium fluxes measured
at EUMELI (values on top, on the left for the oligotrophe site
and the right for the mesotrophe site). We applied François
et al. (1995) algorithm to these fluxes and obtained POC fluxes
(values in green squares) in agreement with results acquired with
other approaches (total primary production, drifting traps, POC
fluxes in steady traps etc…). From Jeandel et al., 2000. Parmi les proxies de productivité les plus fréquemment étudiés (C org., Si biogénique, couple Th/Pa…), la barytine, cristal de baryum de formule BaSO4, est depuis longtemps identifiée comme traceur de la productivité océanique, présente et passée : d’une part, des taux d’accumulation plus importants de barytine ont été observés dans les sédiments des zones de haute productivité (e.g : Brongersma-Sanders, 1967 ; Dehairs et al., 1980 ; Paytan et al., 1993) ; d’autre part une corrélation nette entre les flux de carbone organique et les flux de baryum particulaire, majoritairement présent sous forme de barytine ou de baryum biogène, a été mis en évidence depuis le début des années 80 (Dehairs et al., 1980; Bishop, 1988; Stroobants et al., 1991), avec la publication récente de relations empiriques intéressantes reliant le flux de barytine exporté dans les pièges à particules à la production de surface (Dymond, 1992; Francois et al., 1995; Paytan et al., 1996 ; Jeandel et al., 2000 : un exemple est donné Figure 1).
Le profil du baryum dissous dans l’océan est semblable à celui des nutriments (à ceci près que Ba n’est jamais totalement soustrait en surface, donc jamais considéré comme limitant), indiquant clairement l’influence de l’activité biologique sur le cycle du baryum (Dehairs et al., 1980; Francois et al., 1995; Jeandel et al., 1996). Les travaux de Legeleux et Reyss (1996) montrent que la barytine est formée dans les eaux de surface et sub-surface dans l’Atlantique tropical, eaux pourtant sous-saturées en baryum (Monnin et al, 1999) : ces résultats soutiennent en outre l’hypothèse qu’un processus biologique est à l’origine de la synthèse de la barytine. Cependant, le mécanisme de formation de la barytine reste encore incompris à ce jour. La plupart des auteurs invoquent deux processus possibles : il est évoqué (mais pas démontré) que le phytoplancton est capable de précipiter directement le cristal de baryum ; une autre hypothèse avancée est qu’il y a sursaturation vis-à-vis de la barytine lors de la dégradation des agrégats organiques (Chow et Goldberg, 1960 ; Dehairs et al., 1987 ; Bishop, 1988; Stroobants et al., 1991; Dymond et al., 1992 ; Bertram et al., 1997 ; Jeandel et al., 1996, 2000).
Dans le cas de la première hypothèse, on ne sait pas quelle espèce de phytoplancton accumule préférentiellement la barytine : aucune espèce phytoplanctonique pélagique synthétisant de la barytine n’a pu être isolée jusqu’à présent. L’hypothèse d’une précipitation indirecte par enrichissement en ions sulfates lors de la dégradation de la matière organique dans les pelotes fécales et autres environnements réducteurs est donc privilégiée. Cependant, la concentration en ions sulfates dans l’océan global étant déjà très élevée alors que toutes les eaux de surface sont sous-saturées en Ba (Monnin et al, 1999), l’hypothèse d’un enrichissement en baryum lors de l’activité vivante des organismes, qui serait relibéré lors de la dégradation biologique, semble plus plausible. Bernstein et al. (1992 et 1998), par exemple, suggèrent un apport de baryum dans les agrégats par les acanthaires lors de la dissolution de leur squelette de célestite (SrSO4). Le squelette de ces protozoaires planctoniques marins contient en effet du baryum en concentration environ 10 fois supérieure à celle de l’océan. Une accumulation active du baryum par le phytoplancton pourrait aussi être envisagée. En 1967, suite aux travaux de Vinogradova et Koval’skiy (1962), M. Brongersma-Sanders a avancé l’idée d’une accumulation du baryum par les diatomées pour expliquer son transport vertical. Dehairs et al. (1980), puis Fisher et al. (1991) ont voulu tester l’hypothèse mais n’ont rien observé de significatif. Des expériences d’incubation récentes (Sternberg, 2001) laissent néanmoins penser que les diatomées pourraient fixer le baryum dans leur partie organique, ce que ne feraient pas les coccolithophoridées. Dans le cadre de sa thèse, T. Cattaldo (VUB/LEGOS) a aussi mené des cultures d’algues et suggère que les phaeocystis accumulent plus activement la barytine que d’autres espèces.
Par ailleurs, des travaux ont montré que les rapports isotopiques du radium de la barytine prélevée dans la colonne d’eau pouvaient renseigner sur la profondeur à laquelle ces cristaux se formaient (Legeleux et Reyss, 1996). En effet, les cristaux de barytine incorporent au cours de leur formation les isotopes du radium avec un rapport 228Ra/226Ra égal à celui de l’eau de mer à la même profondeur. En raison de la période relativement courte (5,7 ans) du 228Ra, ce rapport isotopique décroît d’un facteur 20 dans les 500 premiers mètres de la colonne d’eau. Le rapport 228Ra/226Ra peut donc être considéré comme un indicateur relativement sensible de la profondeur. Cependant, si la gamme de profondeur étudiée en 1996 a permis d’affirmer que les cristaux de barytine incorporaient le radium des eaux de surface, elle n’était pas assez détaillée pour que l’on puisse préciser la profondeur de leur formation et par conséquent apporter des informations exploitables en terme de mécanisme de formation. L’effort entrepris par les différentes équipes du projet permettra d’avancer sur ce point.
De plus, la dissolution des particules dans les eaux de fond et à l’interface eau-sédiment est susceptible d’affecter les rapports élémentaires enregistrés par la barytine, comme cela a été suggéré par l’étude du rapport Sr/Ba (Bertram et Cowen, 1997). Moore et Dymond (1991) rapportent également une diminution du rapport 226Ra/Ba mesuré dans la barytine avec la profondeur de la colonne d’eau que ces auteurs ont attribué à un apport latéral de cristaux avec une signature différente. Cependant, il est possible que la dissolution de la barytine puisse affecter également son rapport 226Ra/Ba. Les travaux de P. van Beek menés dans la barytine sédimentaire (van Beek, 2001), suggèrent des variations des rapports 226Ra/Ba (au cours du temps) et Sr/Ba (en fonction de la profondeur de la carotte dans la colonne d’eau) qui nécessitent cependant une meilleure compréhension des processus susceptibles d’affecter ces rapports dans la colonne d’eau. Une partie de ces travaux ont été réalisés dans le cadre du projet BARYTINE (responsable : E. Viollier) soutenu par PROOF en 2000 et 2001 (pas de demande cette année).
En outre, la préservation dans l’enregistrement sédimentaire de la barytine dépend non seulement de son transfert dans la colonne d’eau mais aussi de sa possible dissolution dans les sédiments. Dans certains sédiments de la marge continentale, l’étude des profils de baryum dissous dans les eaux interstitielles, les mesures directes des flux de baryum dissous du sédiment vers la colonne d’eau et l’étude des compositions sédimentaires, révèle une régénération du baryum sous l’interface eau-sédiment (Von Breymann et al., 1992 ; MacManus et al., 1994; Kumar et al., 1996; Paytan and Kastner, 1996, McManus et al., 1998). Cet effet est également observé dans les bassins lacustres où les sédiments sont composés en majorité de débris de diatomées (Viollier, 1995 ; Viollier et al., 1997, Alberic et al., 2000). La source de cette régénération est encore mal connue. La barytine pouvant y contribuer de manière significative, il est important de connaître les facteurs cinétiques qui contrôlent in situ la dissolution/précipitation de ce minéral.
D’une part,
les premières investigations menées sur le comportement
de la barytine dans les sédiments superficiels par E.
Robin et C. Rabouille au LSCE, grâce essentiellement à la
nouvelle technique MEB/EDS, dans le cadre du projet BARYTINE
(Responsable : E. Viollier) semblent indiquer que la barytine
se dissout peu sous l’interface eau-sédiment.
Cette conclusion qui n’est pour le moment qu’une
hypothèse s’appuie sur deux faits essentiels :
(1) la concentration en barytine ne diminue pas en fonction
de la profondeur (elle a même tendance à augmenter
dans les sédiments de l’Atlantique), (2) la distribution
en taille des barytine sédimentaires ne change pas avec
la profondeur dans le sédiment contrairement à ce
qui se passe dans la colonne d’eau où une variation
de cette distribution signe une dissolution.
D’autre part, La concentration en baryum dissous dans les eaux interstitielles
atteint pourtant très rapidement une concentration fixe (250 nmol/L)
qui est au dessus de la concentration à saturation (calculée)
vis à vis de la barytine. Il est même observé une très
nette augmentation de la teneur en Ba dissous dans le premier centimètre
de sédiment.
L’augmentation de Ba dissous et la résistance à la
dissolution du porteur majoritaire du baryum dans les sédiments étudiés
semblent contradictoire. Ce paradoxe peut être interprété,
si une autre phase portant du Baryum (déposée en même
temps que la barytine) se dissout plus vite et sature les eaux interstitielles,
limitant ainsi la dissolution de la barytine. Cette phase solide (minérale
et/ou organique) serait plus abondante à l’interface eau-sédiment
et présente en faible quantité en profondeur.
Comprendre mieux
les processus qui déterminent la synthèse et
la réactivité de la barytine dans la colonne
d’eau et le sédiment de surface est donc important
pour les applications du proxy de productivité : au
delà du lien quantitatif qui pourra être établi
avec la production primaire (et/ou exportée), cela nous
permettra de comprendre quel type de chaîne trophique,
quelles espèces vivantes sont indispensables à la
synthèse de ce cristal. En particulier , le “ rendement ” de
fabrication de la barytine (comparaison entre le rapport POC/bioba
dans le phytoplancton et flux de barytine exportée)
sera un paramètre important à estimer. Sa présence
dans le sédiment pourra alors retracer des taux de production
mais aussi des conditions environnementales nécessaires à cette
production, et à sa préservation.
|
Le principal objectif de cette recherche collective (fédération de 6 laboratoires nationaux, Coll. avec WHOI et Princeton) sur la barytine est de nous faire progresser sur ces questions en couplant à la fois une approche expérimentale menée sur des cultures (Coll. LEGOS-Univ. Princeton, USA), des cinétiques expérimentales (LGE, LSCE) et une approche de terrain (LSCE) qui fait l’objet du projet déposé ici.
|
The goals of this project are : 1) to determine apparent kinetics
2) to identify
3) to confirm (or invalidate) empirical algorithms proposed by François et al. (1995), Dehairs et al. (1997), and Paytan et al. (1996) between barite formation and export production. With this aim, we propose a multidisciplinary approach of barite synthesis in the water column and of its preservation during its vertical transport and in surface sediments. We suggest the determination of the seasonal variability of the barium/barite distribution in dissolved and all accessible particulate fractions (including phytoplankton, zooplankton and aggregates) between the January 2003 and the july 2003, at the DYFAMED site. Our approach consists in coupling a non-destructive observation of the samples (MEB, optical obs. And counting) to chemical analysis. Biological determinations will be conducted carefully, crossing several approaches (for ex. for phytoplankton: species determination using optical microscopy, MEB, via pigments and via flux cytometry). We will combine these measurements to those of 234Th (indicator of export production) and of radium isotopes (identification of the depth to which barite is formed). This project involves six national laboratories and is carried out in collaboration with two international laboratories.
They are all coupled with the SODYF cruises, at the DYFAMED Time Serie Station. |
.
La première publication, réalisée dans le cadre du projet Barytine (PROOF), atteste de notre envie et capacité à travailler ensemble, la seconde est le fruit de la 1ere année de thèse de E Sternberg.
1- Robin E, C. Rabouille, G. Martinez, I. Lefevre, J-L Reyss, P. van Beek
and C. Jeandel. Direct bio-Ba dtermination using SEM/EDS-ACC system
: implication for barite preservation in marine sediments. Submitted
to Marine Chemistry, Juin 2002
2- Sternberg E., Jeandel C. and Morel F.M. Ba uptake by the diatom Thalassioria
Weissflogii: experimental study. Submitted to Marine Chemistry
Références bibliographiques du responsable et de l'équipe (trois dernières années)
Sur
les mesures de paramètres biologiques, travail en
collaboration sur KERFIX
Razouls S., Du Réau G., Guillot P., Maison J. and C. Jeandel. Seasonal
abundance of copepode assemblage and grazing pressure in the Kerguelen
Island area (Southern Ocean) Journal of Plankt. Res ., 20, 1599-1614, 1998.
Ternois Y., M-A Sicre, A. Boireaut, L. Beaufort and C. Jeandel Hydrocarbons,
sterols, and alkenone in sinking particles, at Kerfix site, Southern Ocean.
Org. Geochemistry, 28, 489-501, 1998.
Fiala M., E. Kopczynska, C. Jeandel, L. Oriol & G. Vetion. Seasonal
and interannual variability of size fractionated phytoplankton biomass
and community structure at Station Kerfix, off Kerguelen Islands, Antarctica.
J. Plankt. Res., 20, 1341-1356, 1998.
Sur
le Baryum/barytine
Monnin, C., Jeandel, C., Cattaldo, T. et Dehairs, F., 1999. The marine
barite saturation state of the world's ocean. Marine Chem., 65, 253-261,
1999.
Jeandel C ., Tachikawa K., Bory A. and Dehairs F. Biogenic baryum in suspended
and trapped material as a tracer of export production in the tropical N-E
Atlantic (EUMELI sites). Mar Chemistry, 71, 125-142, 2000
Athias, V., P. Mazzega and C. Jeandel. Estimating the aluminum and baryum
cycling parameters in the tropical NE Atlantic (oligotrophic EUMELI site)
using a genetic algorithm. Journal of Marine Research, in revision, 2001.
Bory A., Jeandel C., Leblond N ; VanGriesheim A., Khripounoff A., Beaufort
L, Rabouille C., Nicolas E ;, Tachikawa K., Etcheber H. and P. Buat-Ménard.
Particle fluxes within different trophic regimes off the Mauritanian upwelling
zone (EUMELI program) Deep Sea Res, in press, 2001
Cattaldo, T., F. Dehairs, N. Metzl, C. Monnin and C.Jeandel Hydrological
and Biogeochemical Control of Dissolved Baryum in the Southern Ocean along
6 degrees W and 30 degrees E, Journ. Geoph. Res. En révision
Sternberg E., Accumulation of baryum by marine phytoplankton, Mémoire
DEA “Océanologie, météorologie, environnement ” Univ
Paris VI, 30p, 2001
P. Albéric, E. Viollier, D. Jézéquel, C. Grosbois and G. Michard (2000), Trace elements-dissolved organic matter interaction in the anoxic deep layer of Lake Pavin (Puy-de-Dôme, France), Limnology & Oceanography, 45(5), pp1088–1096
Associé au
projet « BARYTINE »
E. Robin, C. Rabouille, G. Martinez, I. Lefevre, C.
Jeandel, P. Van Beek and J-L. Reyss, Barite in marine sediments:
a comparison between a new SEM/EDS technique with ICP-MS and
INAA methods, in prep. pour Marine Chem.
P. van Beek, J.-L. Reyss, R. Gersonde, M. Paterne, M. Rutgers van der Loeff, G. Kuhn, 226Ra in barite : Absolute dating of Holocene Southern Ocean sediments and paleoceanographic implications, soumis à Journ. Geoph. Res.
Van Beek P. and Reyss J.-L. (2001) , 226Ra in marine barite : new constraints on supported 226Ra, Earth and Plan. Sci. Let., 187, 147-161
Van Beek P., 2001. Utilisation de la barytine sédimentaire marine pour les études paléocéanographiques de l’Holocène, Thèse, Paris VI, 179pp.
NB : le
travail ci-dessous est aussi une production “ EIMETO ”
Arraes-Mescoff R., M. Roy Barman, L. Coppola, M. Souhaut, K. Tachikawa,
C. Jeandel, R. Sempéré and C. Seri-Yoro, The behavior of
Al, Mn, Ba, Sr, REE and Th isotopes during in vitro degradation of large
marine particles Marine Chemistry, 73, 1-19, 2001.
Sur
le PVM
Gorsky G., Picheral, M. and Stemmann, L. (2000a) Use
of the Underwater Video Profiler for the Study of Aggregate Dynamics
in the North Mediterranean. Estuar. Coast. Shelf Sci. 50 (1)
: 121-128.
Stemmann L., M. Picheral & G. Gorsky (2000) Diel changes in the vertical distribution of suspended particulate matter in the NW Mediterranean Sea investigated with the Underwater Video Profiler. Deep Sea Res. I, 47 : 505-531.
Sur
le 234Th
Hamilton, T.F., J. Heilmann, S.W Fowler and J.C. Miquel,
(1999). Spatial variations of scavenging rates within the Middle
and Northern Adriatic Sea: use of U-Th disequilibria. . In: Hopkins,
T.S., A. Artegiani, G. Cauwet, D. Degobbis and A. Malij (eds.),
Physical and Biogeochemical Processes in the Adriatic Sea, Ecosystem
Research Report 32 The Adriatic Sea, EUR 18834, Directorate-General
for Research, European Commission, Brussels, 525-535.
Miquel, J.C, (1999). Hydrothermal Fluxes and Biological Production in the Aegean Sea: Large Volume in-situ Sampling of Seawater. In: Hieke, W., C. Hemleben, P. Linke and H. Weikert, MITTELMEER 97/98, Reise Nr.40, 28. Oktober 1997 - 10.Februar 1998, Meteor-Berichte, Universitat Hamburg, 99-X. pp.32-34.
Miquel, J.-C., E.N. Laurent, S.W. Fowler and K. Cochran, (2001). Carbon Export in Coastal NW Mediterranean Sea: Sediment Trap Records and 234Th:238U Disequilibria Modelling. Eos Trans. AGU, 82(47), Fall Meet. Suppl., Abstract OS31C-0459.
Sur la cinétique expérimentale
Roychoudhury A., Viollier E. and Van Cappellen P. (1998), A plug-flow reactor for studying biogeochemical reactions in undisturbed aquatic sediments, App. Geochem., 13, 269-280
Autres références citées :
Bernstein, R.E., Byrne, R.H., Betzer, P.R. and Greco, A.M., Morphologies and transformations of celestite in seawater: The role of acantharians in strontium and baryum geochemistry, Geochimica et Cosmochimica Acta, 56, 3273-3279, 1992.
Bernstein, R.E., Byrne, R.H., Schijf, J., Acantharians: a missing link in the oceanic biogeochemistry of baryum, Deep-Sea Research I, 45, 491-505, 1998.
Bertram, M.A. and Cowen, J.P., Morphological and compositional evidence for biotic precipitation of marine barite, Journal of Marine Research, 55 (3), 577-593, 1997.
Bishop, J.K.B., The barite-opal-organic carbon association in oceanic particulate matter, Nature, 332, 341-343, 1988.
Brongersma-Sanders, M., Baryum in pelagic sediments and in diatoms, Proceedings, Series B, 70 (1), 93-99, 1967.
Chow, T.J. and Goldberg, E.D., On the marine geochemistry of baryum, Geochimica et Cosmochimica Acta, 20, 192-198, 1960.
Dehairs, F., Chesselet, R. and Jedwab, J., Discrete suspended particles of barite and the baryum cycle in the open ocean, Earth and Planetary Science Letters, 49, 528-550, 1980.
Dehairs, F., Lambert, C.E., Chesselet, R., and Risler, N., The biological production of marine suspended barite and the baryum cycle in the western Mediterranean Sea, Biogeochemistry, 4, 119-139, 1987.
Dymond, J., Suess, E. and Lyle, M., Baryum in deep-sea sediment: a geochemical proxy for paleoproductivity, Paleoceanography, 7 (2), 163-181, 1992.
Fisher, N.S., Guillard, R.R.L. and Bankston, D.C., The accumulation of baryum by marine phytoplankton grown in culture, Journal of Marine Research, 49, 339-354, 1991.
Francois, R., Honjo, S., Manganini, S.J. and Ravizza, G.E., Biogenic baryum fluxes to the deep sea: Implications for paleoproductivity reconstruction, Global Biogeochemical cycles, 9 (2), 289-303, 1995.
Jeandel, C., Dupré, B., Lebaron, G., Monnin, C. and Minster, J.-F., Longitudinal distributions of dissolved baryum, silica and alkalinity in the western and southern Indian Ocean, Deep-Sea Research I, 43 (10), 1-31, 1996.
Legeleux, F. and Reyss, J.-L., Activity ratio in oceanic settling particles: Implications regarding the use of baryum as a proxy for paleoproductivity reconstruction, Deep-Sea Research I, 43 (11-12), 1857-1853, 1996.
Paytan, A., Kastner, M., Martin, E.E., Macdougall, J.D. and Herbert, T., Marine barite as a monitor of seawater strontium isotope composition, Nature, 366, 445-448, 1993.
Paytan, A., Kastner, M. and Chavez, F.P., Glacial to Interglacial Fluctuations in Productivity in the Equatorial Pacific as Indicated by Marine Barite, Science, 274, 1355-1357, 1996.
Stroobants, N., Dehairs, F., Goeyens, L., Vanderheijden, N. and Van Grieken, R., Barite formation in the Southern Ocean water column, Marine Chemistry, 35, 411-421, 1991.
Vinogradova, Z.A. and Koval’skiy, V.V., Elemental composition of Black Sea plankton, Dokl. Akad. Nauk SSSR, earth sc. sect., 147, 217-219, 1962.
Bertram M.A. & Cowen J.P., 1997. Morphological and compositional evidence for biotic precipitation of marine barite, J. Mar. Res. 55, 577-593
Moore W.S. & Dymond J., 1991. Fluxes of Ra-226 and barium in the Pacific Ocean : The importance of boundary processes, Earth Planet. Sci. Lett. 107, 55-68
Paytan A., Kastner M. & Chavez F.P., 1996a. Glacial to Interglacial fluctuations in productivity in the equatorial Pacific as indicated by marine barite, Science 274, 1355-1357
Paytan A, Moore W.S. & Kastner M., 1996b. Sedimentation rate as determined by 226Ra activity in marine barite, Geochim. Cosmochim. Acta 60 (22), 4313-4319
Klinkhammer G.P. and Chan L.H. (1990), Determination of barium in marine
waters by isotope dilution inductively coupled mass spectrometry, Anal.
Chim. Acta, 232, 323-329
Kumar, N., Anderson, R.F. and Biscaye, P.E. (1996), Remineralization of particulate authigenic trace metals in the Middle Atlantic Bight: Implications for proxies of export production, Geochim. Cosmochim. Acta, 60, 3383-3397
MacManus J., Berelson W.M., Klinkhammer G.P., Kilgore T.E. and Hammond D.E. (1994), Remobilization of barium in continental margin sediments, Geochim. Cosmochim. Acta, 58, 4899-4907
McManus J., Berelson W.M., Klinkhammer G.P., Johnson K.S., Coale K.H., Anderson R.F., Kumar N., Burdige D.J., Hammond D.E., Brumsack H.J., McCorkle D.C., Rushdi A. (1998), Geochemistry of barium in marine sediments : implications for its use as paleoproxy, Geochim. Cosmochim. Acta, 21-22, 3453-3473
Van Cappellen P. and Qiu L. (1997), Biogenic silica dissolution in sediments of the Southern Ocean. I. Solubility, Deep-Sea res. II, 44, 1109-1128
Viollier E. (1995) Géochimie des éléments traces en milieu lacustre, Thèse de l’Université Paris 7, 330 p
Viollier E., Michard, P.Alberic, D. Jezequel, F. Prevot and G. Sarazin (1997) Geochemical study of a crater lake : Lake Pavin, Puy de Dôme, France - Constraints afforded by the particulate matter distribution in the element cycling within lake, Chem. Geol., 142, 225-241
Von Breymann
M.T., Emeis K.C. and Suess E. (1992), Water depth and diagenetic
constraints on the use of barium as a paleoproductivity indicator,
In Upwelling Systems : Evolution Since the Early Miocene, Geol.
Soc., London, special publication 64, 273-284




